
記住我
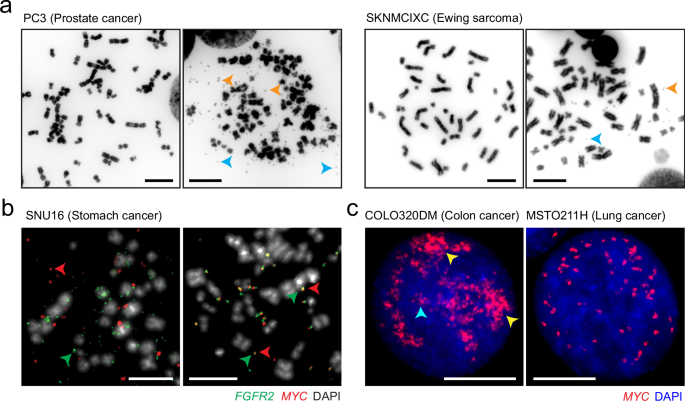
ecDNA is prevalent in human cancer. The first systematic pan-cancer investigation has estimated that ecDNA is present in nearly half of all cancer types and up to one-third of all cancer patients throughout their disease, while it was not found in matched blood and adjacent tissues.4 This study also revealed that ecDNA is associated with shorter survival across multiple cancers, even after adjusting for tissue type.4 Subsequent studies focused on specific cancer types, such as small cell lung cancer,27 neuroblastoma,61 medulloblastoma,26 and hepatocellular carcinoma,54 have similarly demonstrated that ecDNA is linked to poorer clinical outcomes.
A debate has since been raised: Is ecDNA a genuine cancer driver promoting early malignant transformation, or is it just a manifestation in late-stage cancer due to genomic instability? This appears to be a “chicken-and-egg” dilemma, but it turns out that both arguments are true (Fig. 5). As previously discussed, a longitudinal, case-control study in BE and associated EAC has shown that ecDNAs occur in the pre-cancerous lesions that eventually progressed to cancer.7 More importantly, ecDNAs were not found in pre-cancerous lesions that did not transform, even with long-term follow-up.7 Although further prospective studies are still required to clarify the causality between ecDNA and malignant transformation, it has provided strong evidence showing that ecDNAs can be generated preceding tumor formation. Another piece of evidence comes from a preprint study modeling ecDNA formation via the Cre-loxP system.62 The Cre recombinase can induce the circularization of a DNA segment flanked by two loxP sites with the same orientation. It has been shown that Cre-loxP-generated MDM2 ecDNA enables malignant transformation in mouse embryonic fibroblasts driven by over-expression of the HRAS G12V mutant.62 While it has not been directly demonstrated that ecDNA alone is sufficient to drive transformation, applying this technology to generate specific ecDNAs in an appropriate tissue and developmental context is a promising way to address this question.
Fig. 5: ecDNA in tumor formation, evolution, and drug resistance.
ecDNAs may emerge before tumor formation, and subsequently drive malignant transformation. Along with tumor evolution, ecDNA copy number, complexity, and heterogeneity can increase. Under selective pressure of therapies, specific subpopulations of tumor cells with optimal ecDNA composition survive, rendering drug resistance and relapse.
The most well-known challenge brought by ecDNA is therapy resistance. This was first documented nearly half a century ago when it was discovered that ecDNA amplification of the DHFR gene is responsible for methotrexate resistance.63,64 Methotrexate is an antimetabolite of folic acid that binds DHFR to inhibit the conversion of dihydrofolate into tetrahydrofolate, which is required for nucleotide synthesis. As a potent gene amplification mechanism, ecDNA can raise the DHFR copy number to an enormously high level, which, in turn, produces a massive amount of DHFR protein to dilute the effect of methotrexate. However, the rapid fluctuation in ecDNA copy number presents an even greater challenge. ecDNAs segregate randomly and create a heterogeneous pool of cancer cells containing diverse ecDNA species and subspecies of various copy numbers. This rich repertoire of heterogeneity provides adaptive fitness under the selective pressure of therapy.65
In certain contexts where high oncogenic output is beneficial, cancer cells with high ecDNA copy numbers are selected (Fig. 5). One of the most well-articulated examples is the acquired cross-resistance in small cell lung cancer driven by ecDNAs with MYC family paralogs.37 While most small cell lung cancer cases are initially chemosensitive, they often develop cross-resistance to multiple chemotherapy regimens upon relapse.66,67 A study comparing patient-derived xenografts derived before and after treatment has shown that chemotherapy can induce the formation of ecDNAs, including those carrying MYC.37 When chemoresistant xenografts are challenged by chemotherapies, DNA damage is selectively observed in the tumor cell population with low MYC ecDNAs. Consequently, tumor cells with high MYC ecDNAs are selected, leading to an increase in the overall MYC copy number over time.37 This study provides a direct, single-cell observation of the adaptive advantage conferred by ecDNA heterogeneity. A similar observation has been made in melanoma patients, in which mitogen-activated protein kinase (MAPK) inhibitors select for ecDNAs carrying resistance genes, such as BRAF.24
Conversely, tumor cell populations with low ecDNAs may be advantageous under different therapy pressures, especially with targeted therapy (Fig. 5). While clinical evidence is currently limited, experimental data using glioblastoma patient-derived cells have shown that the epidermal growth factor receptor (EGFR) kinase inhibitor erlotinib can deplete EGFR ecDNAs.21 Once the targeted drug loses its target, cancer cells become resistant. However, upon drug withdrawal, EGFR ecDNAs rapidly re-emerge within a few cell doublings.21 Given the fast kinetics of ecDNAs and their remarkable contributions to therapy resistance, it will be necessary to monitor ecDNA dynamics in clinical practice.
The understanding of ecDNA has been expanded with the discovery that ecDNA may carry immunoregulatory genes, such as PD-L1 in HPV fusion ecDNA and those identified in BE.7,8 This has sparked a growing interest in exploring how ecDNA-driven cancer may interact with the tumor microenvironment. A transcriptomic study based on bulk tumor RNA sequencing from The Cancer Genome Atlas (TCGA) suggests that cancers bearing ecDNA may exhibit an “immune-cold” phenotype. This is characterized by reduced immune cell infiltration and lower expression of antigen-presenting molecules compared to ecDNA-negative tumors.68 In line with this, another study using a machine-learning approach on transcriptomic data from TCGA has similarly discovered that ecDNA-positive cancers downregulate immunomodulatory processes, including lymphocyte activation, leukocyte chemotaxis, and cytokine production, which are integral to many facets of the cancer immune response.69
However, our knowledge of the detailed immunosuppressive landscape in ecDNA-positive cancers and the exact mechanism that drives the immune-cold phenotype is still limited. Studies so far are based on bulk-cell sequencing data, making it difficult to deconvolute cell composition and crosstalk within the tumor microenvironment. Furthermore, experimental interrogation is lacking; therefore, the causality between the ecDNA-positive tumors and the immune-cold phenotype is still unclear. More specifically, it is unclear whether ecDNA-bearing tumors can actively shape their tumor microenvironment or whether the immune-cold microenvironment favors the formation of ecDNAs in cancer cells. Applying single-cell and spatial genomic technologies to ecDNA mouse tumor models will open a new opportunity to understand the interplay between tumors with ecDNA and their microenvironment. This could potentially lead to significant advancements in our understanding of ecDNA biology, particularly its role in tumor development and progression, as well as provide new insights into devising immunotherapy approaches for ecDNA-driven cancers.
Emerging thoughts on ecDNA-directed therapyecDNA has become a novel target for cancer therapy due to its unique role in cancer biology. First, ecDNA has shown pivotal roles in cancer development and therapy resistance. Second, ecDNA is exclusively present in cancerous and pre-cancerous but not in normal tissues. Third, targeting the protein products encoded by ecDNAs has proven challenging due to the dynamic nature and heterogeneity of ecDNA.15,21 More compellingly, targeting the oncogene-carrying ecDNA will make many currently undruggable targets actionable. While experimentally depleting ecDNA via CRISPR cutting is possible in cell culture,15 it is hard to imagine translating this approach into gene therapy for patients, considering the ongoing structural evolution of ecDNA in tumors. Current research endeavors focus on attacking ecDNA during both its formation and maintenance.
Targeting ecDNA formationMutational signature analysis on paired patient-derived xenografts, both before and after treatment, has unequivocally demonstrated that chemotherapy can induce the formation of ecDNAs and foster resistance.37 This finding prompts the question of whether the inhibition of ecDNA formation could serve as a feasible therapeutic option. Therefore, a comprehensive understanding of the pathogenesis mechanism of ecDNA is essential for effective prevention of ecDNA formation.
In theory, ecDNA formation requires the initial breaks in its chromosome ancestors, resulting in the release of DNA segments that are subsequently re-ligated into a circular form. This process of cut-and-re-ligation explains why the presence of ecDNAs is often associated with deletion events in the original chromosomes (Fig. 6a). This phenomenon has been documented via southern blot, FISH, and sequencing in multiple cancer types.70,71,72 Experimental modeling of DNA double-strand breaks has confirmed that it is mechanistically possible. CRISPR-liberated chromosomal DNA segments can be self-ligated and form circular ecDNA, which can then be selected under appropriate selection pressure.15,73
Fig. 6: Scar-prone and scar-free ecDNA formation.
a ecDNA formation with a scar on the chromosome. A chromosome undergoes mild DNA damage, such as two DNA double-strand breaks within a certain distance, releasing a linear DNA fragment. The broken chromosome and the free DNA fragment are re-ligated via end-joining individually, forming a circular ecDNA and a repaired chromosome with a focal deletion (scar). b ecDNA formation with multiple scars on the chromosome. A chromosome undergoes pulverization during chromothripsis, resulting in many DNA fragments. Some fragments ligate with each other to form ecDNA, while others reassemble into a heavily rearranged chromosome with multiple scars. c ecDNA formation without a scar on the chromosome. If mild DNA damage occurs in the replication forks during the S phase, the broken chromosome can be repaired by homologous recombination, leaving no scar on the chromosome while an ecDNA forms. d Another possible way to generate ecDNA without leaving a scar on the chromosome. Whole-chromosome or whole-genome duplication occurs before one of the chromosomes shatters. While the broken chromosome is depleted, ecDNA can arise from the fragmentized DNA, resulting in no visible scar on the chromosome.
While CRISPR models ecDNA formation from a mild focal deletion, ecDNA can also originate from more severe genomic instability contexts. For example, ecDNAs can arise from chromothripsis, a process where a chromosome mis-segregates and shatters into small pieces through multiple mechanisms, such as TREX1-mediated DNA resection,74 as well as pathological DNA damage responses via base excision repair75 and Fanconi anemia pathway.76 The shattered pieces are then re-ligated through DNA repair (such as non-homologous end-joining).77,78 During the re-ligation, centromere-less DNA fragments may be circularized to form ecDNA, resulting in complex rearrangements on both the derivative chromosome and ecDNA (Fig. 6b).22,77,79 In a pan-cancer study using the TCGA dataset, signatures of chromothripsis were detected in 36% of ecDNA amplicons and over 50% of samples with ecDNA,4 highlighting a significant role of chromothripsis in ecDNA formation.
However, certain studies have also shown that the presence of ecDNA does not necessarily correlate with a loss of heterozygosity.72,80 This observation suggests a copy-and-re-ligation hypothesis, in which DNA damage occurs in a replication fork, and the focally resected DNA scar is repaired by homologous recombination (Fig. 6c).80 However, although mechanistically possible, it still lacks evidence to support that the copy-and-re-ligation process naturally occurs and gives rise to ecDNA.
Alternatively, the cut-and-re-ligation model can still explain the scar-less ecDNA formation. If a whole-chromosome or whole-genome duplication event precedes the focal deletion, and the broken chromosome is then depleted due to processes like mis-segregation, the final outcome will appear as if there is no scar on the chromosome (Fig. 6d). This hypothesis is quite plausible because the formation of ecDNA is associated with the loss of function of TP53.7,69 A longitudinal study in BE and associated EAC shows that TP53 deficiency precedes the formation of ecDNA.7 A TCGA pan-cancer analysis further shows that TP53 is the only gene whose mutations are significantly higher in ecDNA-containing tumors.69 Therefore, when the compromised TP53 primes genomic instability, such as whole-genome doubling,81 scar-less ecDNA formation is possible under the cut-and-re-ligation context.
After DNA segments are liberated from chromosomes, how are they re-ligated to form ecDNA? Breakpoint analyses suggest the existence of multiple possible pathways, including classic and alternative non-homologous end-joining and homologous recombination, with frequency varying among cancer types and treatment conditions.72,80,82 In line with these studies, inhibition of DNA-PKcs or PARP, both of which are important regulators of DNA repair pathways, can decrease the frequency of ecDNA formation under the context of chromothripsis.22,82,83 However, it remains an open question as to whether ecDNA formation can bypass such inhibition via alternative pathways. Although targeting multiple pathways is possible, we posit that any strategies aimed at targeting ecDNA formation must be carefully tailored to the specific cancer type and treatment context to mitigate the risk of toxicity from unnecessary medical interventions.
Targeting ecDNA maintenanceWhen a tumor is already ecDNA-positive at diagnosis, it is more rational to eliminate ecDNA rather than prevent ecDNA formation for therapy. If critical mechanisms of ecDNA maintenance are disrupted, it is possible to deplete ecDNA in cancer.
How is ecDNA maintained within the cancer genome? Similar to chromosomal DNA, ecDNA relies on at least three essential processes for their maintenance: (1) replication before mitosis to maintain their copy number; (2) segregation to ensure inheritance among daughter cells; (3) repair upon DNA damage. Additionally, genomic stability is overseen by the cell cycle checkpoint and innate immunity surveillance.84,85,86 Given that ecDNA is a manifestation of genomic instability, there is growing interest in understanding how ecDNA may evade the checkpoints of the cell cycle and innate immunity for their maintenance.
Certain small circular DNA species, such as plasmids and bacteriophage genomes, can duplicate through rolling-circle replication (RCR).87 However, chromatin-containing ecDNA of megabase sizes in cancer is unlikely to be replicated by RCR, because mammalian cells lack essential genes to initiate RCR, such as RepB and RepC. Early investigation using Okazaki fragment hybridization mapping indicated that ecDNA contains a bidirectional replication origin.88 In addition, differential staining of sister chromatids using thymidine analogs has shown that ecDNA replicates once and only once during the S phase in a semi-conservative manner, showing double-minute morphology.29,89 These observations suggest that ecDNA replication may employ machinery similar to that used for chromosomal DNA. Consequently, the opportunity to target ecDNA may not lie in the DNA synthesis process itself, but rather in how ecDNA and chromosomal DNA mitigate replication stress, which is a common feature in cancer.90
DNA replication, while it might seem like a simple “copy-paste” process, often encounters many obstacles, such as DNA secondary structures, DNA–RNA hybrids, and limited nucleotide availability.91 These obstacles lead to replication stress, a common feature of the cancer genome.90 It has been proposed that ecDNA is more susceptible to replication stress, because hydroxyurea, a replication stress inducer that inhibits ribonucleotide reductases to reduce the nucleotide pool, can induce ecDNA loss in cell culture.92,93 However, it remains unclear whether ecDNA particles are more sensitive to nucleotide shortage, or whether the presence of ecDNA renders cancer cells more vulnerable to hydroxyurea. A recent study argues that ecDNA suffers from a higher degree of replication-transcription conflict due to their elevated transcriptional activity.94 This, in turn, makes ecDNA-bearing cancer cells more vulnerable to the inhibition of CHEK1, a critical checkpoint protein in DNA damage response and cell cycle.94,95,96
Other aspects of the ecDNA maintenance mechanisms are largely unexplored. A TCGA pan-cancer transcriptomic study reveals specific upregulation of DNA double-break repair pathways in ecDNA-bearing tumors, including classic and alternative end-joining, single-strand annealing, and homology-directed repair pathways.69 While this finding has not been experimentally validated, it highlights the importance of understanding how ecDNAs are repaired in different damage contexts and whether there is an ecDNA-specific repair mechanism that could be exploited for pharmacological targeting. Previously, several studies showed that treatments with DNA-damaging agents, such as hydroxyurea and radiation, can lower ecDNA levels.97,98 These findings suggest the potential feasibility of targeting the mechanisms involved in ecDNA damage repair and maintenance. However, because these studies induced DNA damage globally that was not specific to ecDNA, it is challenging to deconvolute ecDNA-specific DNA damage response.
Mechanistic insights into ecDNA segregation also remain limited. While ecDNA has been observed to “hitchhike” on mitotic chromosomes to segregate into daughter nuclei,15,48 the factors mediating ecDNA–chromosome interaction during mitosis have not been identified. Theoretically, the contact (but not hybridization) between double-strand DNA molecules requires protein intermediaries, as the negative charges from the phosphate groups on the DNA backbone would otherwise repel such an interaction; unless DNA and histone modifications (such as methylation and acetylation, which are also protein-mediated processes) happen to create a favorable electrostatic interaction environment perfectly. Early studies on viral episomes, which are circular, extrachromosomal viral DNAs that attach to mitotic chromosomes for segregation, have identified many proteins coded by the virus genome involved in the attachment of episomes to mitotic chromosomes.99,100,101,102 These viral proteins bind to episomes and interact with host proteins on mitotic chromosomes, enabling episome segregation. However, most ecDNAs do not encode any foreign proteins other than those found in the cancer genome (except for the possibility of ecDNA–virus hybrids), it remains unclear what proteins are recruited to mediate the interaction between ecDNAs and mitotic chromosomes. Identifying these factors will provide a better understanding of ecDNA segregation and offer new therapeutic targets.
Research into ecDNA-specific targeting is still in its infant stage, and emerging new ecDNA targeting strategies beyond tackling ecDNA formation and maintenance, such as targeting ecDNA’s mobility, have been proposed.103 We anticipate that it will become one of the most important directions in the next decade. While advances in modeling and screening technologies have facilitated the study of ecDNA-specific maintenance mechanisms, we should be cautious when translating our knowledge yielded from basic science research into clinically actionable targets and trials. Cancer therapy is always a trade-off between efficacy and toxicity. This principle is equally applicable to ecDNA-directed therapy. The degree to which a target or a mechanism is specific to ecDNAs, as opposed to chromosomes, will be a critical factor in determining the therapeutic window.
Clinical diagnosis for ecDNA: Where are we now?With growing evidence supporting that ecDNA can serve as a predictive biomarker for clinical outcomes, such as malignant transformation, patient survival, and therapy response as discussed above, and ecDNA-directed clinical trials are on the horizon, there is a pressing need for the development of more cost-effective tools for ecDNA detection.
In our previous review of the current ecDNA detection toolbox, we highlighted that direct visualization of ecDNAs in cells at metaphase is considered the gold standard.3 The development of computer-assisted image analysis software, such as ECdetect14 and ecSeg,104 has facilitated automatic ecDNA detection. Even when metaphase chromosome preparation is not available, such as in formalin-fixed paraffin-embedded tissue samples,105 there are ongoing efforts to use machine learning technologies to detect ecDNAs by recognizing the signal pattern from FISH in interphase nuclei. Given that cytogenetics has been a standard clinical practice for decades, the imaging-based method is one of the most promising routes to be integrated into the existing clinical pipeline. However, as we have previously discussed, this approach suffers from low throughput, necessitating the development of more advanced, high-throughput technologies. Furthermore, the selection of a FISH probe requires prior sequence information, unless recurrent ecDNAs that contribute to a specific clinical outcome have been identified, such as those amplifying MYC and paralogs in chemoresistant small cell lung cancer.37
In comparison, sequencing-based methods offer a higher throughput. Currently, paired-end whole-genome short-read sequencing remains the most accessible method, as it does not require any special modifications of the experimental protocol for ecDNA detection. The ecDNA information is retrieved by computational analysis using the AmpliconArchitect software, which can analyze the structure of an amplicon by interval search, structural variation detection, and breakpoint graph construction and visualization.106 AmpliconArchitect is the only tool so far to extract ecDNA information from short-read WGS. As such, WGS is more feasible for automation, from sample processing to data interpretation. However, AmpliconArchitect cannot distinguish between ecDNA and ecDNA-originated homogeneously staining regions derived from ecDNA reintegration to the chromosome. The reintegration often maintains the fine structure of multiple copies of ecDNA. Therefore, it is challenging to find a sufficient number of reads to support reintegration junctions in short-read sequencing.107 And while CIRCLE-seq is designed to enrich circular DNA for sequencing by digesting linear DNA,55,
留言 (0)