
記住我
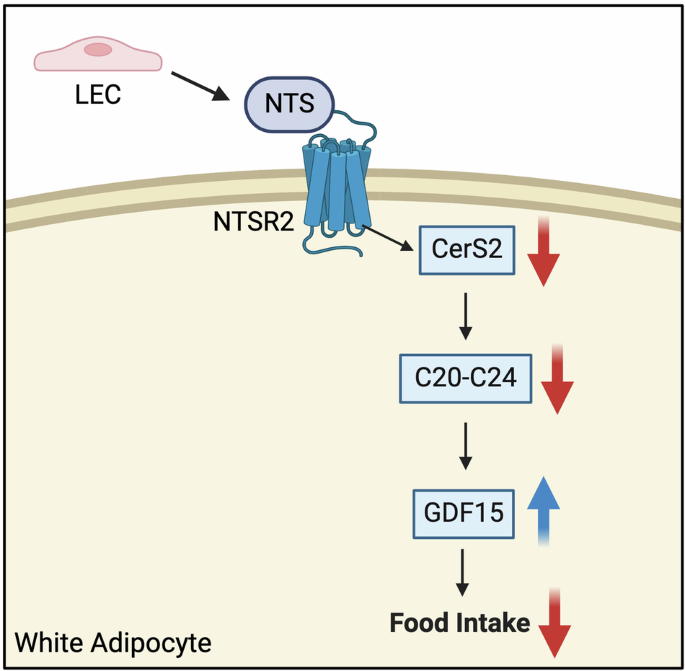
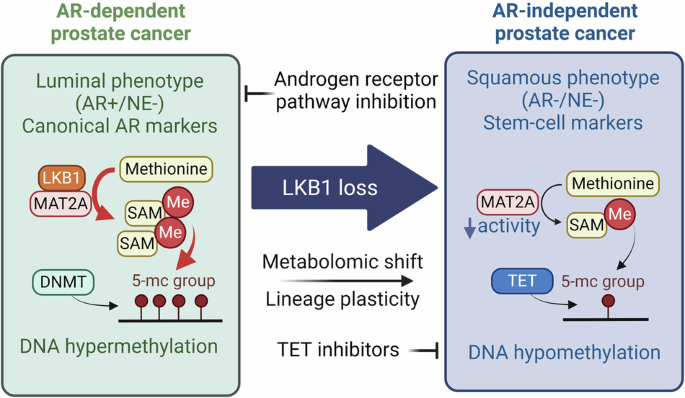
Heat acclimation (HA) is a key adaptive response in mammals to repeated heat exposure, essential for fitness and survival.1,2 HA improves cardiovascular function, thermal comfort, and exercise capacity.3,4 However, the lack of a genetically tractable model has hindered understanding of the molecular and neural mechanisms underlying HA. Here, we show that 10 days of daily 38 °C exposure lowers core body temperature (Tcore) and reduces anxiety during subsequent heat exposures in mice. HA increases brain-derived neurotrophic factor (BDNF) expression in the medial preoptic area (MPO). BDNF-expressing MPO (MPOBDNF) neurons show increased intrinsic heat sensitivity after HA. These neurons orchestrate downstream targets in the dorsomedial hypothalamus (DMH) and rostral raphe pallidus (rRPa) to mediate HA effects. BDNF, acting through its receptor tropomyosin-related kinase B (TrkB) in the DMH, facilitates the anxiolytic effect of HA by enhancing excitatory synaptic connections between MPOBDNF and DMH neurons. This study provides new insights into HA mechanisms, setting the stage for future research on heat stress reduction and exercise optimization.
Without HA, heat exposure (38 °C) increased anxiety but did not impair novel object recognition in male C57 mice, as assessed in the open field test (OFT) and Y-maze test (Supplementary information, Fig. S1a, b). To establish an effective HA protocol, we evaluated three different protocols. Neither daily exposures to 35 °C for 3 h over 14 days nor daily exposures to 35–38 °C for 3 h over 20 days impacted Tcore during subsequent 38 °C exposures (Supplementary information, Fig. S1c, d). However, consecutive 10-day exposures to 38 °C for 2 h significantly improved heat tolerance, as marked by a moderate rise in Tcore during subsequent 38 °C exposures (Supplementary information, Fig. S1e). Thus, we established an effective HA protocol. Under this protocol, food intake decreased while water intake significantly increased during the 2-h heat exposure. Over the full 10-day acclimation period, water consumption increased significantly, while no changes were observed in body weight or food intake (Supplementary information, Fig. S1f–l).
After HA, we re-exposed the mice to 38–40 °C to systematically assess the adaptive effects (Fig. 1a). HA mice displayed extended Tcore tolerance in the heat tolerance test (HTT) at 40 °C, which lasted three times longer than that in the non-HA group, as measured by the latency for Tcore to reach 41.5 °C (a critical thermal limit5) (Fig. 1b). In addition, HA mice exhibited increased open arm exploration in the elevated plus maze (EPM) (Fig. 1c) and central exploration during OFT (Supplementary information, Fig. S2a, b), indicative of reduced anxiety. However, HA did not influence thermal preference or nociceptive heat perception (Supplementary information, Fig. S2c, d). While physical activity remained constant during heat exposure at 38 °C, energy expenditure (EE) reduced (Supplementary information, Fig. S2e, f). This reduction in EE aligned with decreased mRNA levels of uncoupling protein 1 (UCP1) in brown adipose tissue (Supplementary information, Fig. S2g). HA did not affect the time spent or the number of entries into the closed arms in the Y-maze (Supplementary information, Fig. S2h). HA also enhanced thermoregulatory behaviors, as characterized by longer grooming time (Supplementary information, Fig. S2i) and faster, longer body postural extension during heat exposures (Supplementary information, Fig. S2j). Similarly, HA training in female mice also enhanced Tcore tolerance and reduced heat-induced anxiety (Supplementary information, Fig. S3a–c). Thus, HA significantly improves heat tolerance and alleviates anxiety by reducing thermogenesis and bolstering thermoregulatory capacity, without affecting cognitive and pain perception functions.
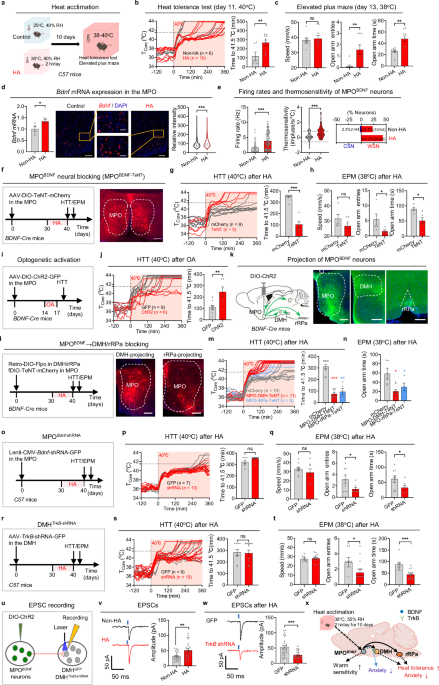
Fig. 1: BDNF in the MPO is required for heat acclimation.
a Experimental procedure for HA. Mice were subjected to controlled conditions of 25 °C and 40% relative humidity (RH) in an incubator. The HA group underwent a 10-day training program consisting of daily 2-h sessions under conditions of 38 °C and 60% RH. b Tcore changes in exposure to ambient temperature of 40 °C in the HTT from individual mice. The time taken for Tcore to reach 41.5 °C was quantified on the right. c EPM under heat exposure (38 °C). The traveling speed, entries into open arm and time spent in open arm were quantified. d Expression of Bdnf mRNA in the MPO in response to a 2-h heat exposure (38 °C), determined by qPCR and RNAscope. The Bdnf mRNA expression of each neuron is quantified on the right. Scale bars, 400 μm (aside) and 100 μm (middle). e Basal firing rates at 36 °C, the thermosensitivity of MPOBDNF neurons and the percentage of warm-sensitive neurons (WSNs) or cold-sensitive neurons (CSNs) within recorded BDNF+ neurons. f Protocol for HA training after blocking MPOBDNF neurons with TeNT and the representative expression of AAV-DIO-TeNT-mCherry. AAV-DIO-TeNT was injected into the MPO of the BDNF-IRES-Cre mice (termed as MPOBDNF-TeNT, TeNT group). Scale bar, 200 μm. g, h HTT (g) and EPM (h) after HA training in MPOBDNF-TeNT mice. i Experimental design to test the effects of optogenetic activation (OA) of MPOBDNF neurons. AAV-DIO-ChR2-GFP was injected into the MPO of the BDNF-IRES-Cre mice (termed as MPOBDNF-ChR2, ChR2 group). j HTT of the MPOBDNF-ChR2 mice after OA. k Axonal GFP expression of MPOBDNF-ChR2-GFP neurons in the DMH and rRPa. Scale bars, 200 μm. l Protocol for HA training after blocking DMH/rRPa-projecting MPOBDNF neurons and representative expression of AAV-fDIO-TeNT-mCherry. Retro-DIO-Flpo was injected into the DMH/rRPa and AAV-fDIO-TeNT-mCherry was injected into the MPO of the BDNF-IRES-Cre mice (MPO-DMH/rRPa TeNT group). Scale bars, 200 μm. m, n HTT (m) and EPM (n) after HA training in MPO-DMH/rRPa TeNT group. o Experimental design to knock down Bdnf in MPO by injecting Lenti-CMV-Bdnf-shRNA into C57 mice (termed as MPOBdnf-shRNA, shRNA group). p, q HTT (p) and EPM (q) after HA training in MPOBdnf-shRNA mice. r Experimental design to knock down TrkB in DMH neurons by injecting AAV-hSyn-Cre and AAV-DIO-TrkB-shRNA-GFP into C57 mice (termed as DMHTrkB-shRNA, shRNA group). s, t HTT (s) and EPM (t) after HA training in DMHTrkB-shRNA mice. u Experimental design to record postsynaptic currents in DMH neurons innervated by MPOBDNF neurons using whole-cell patch configurations. v Representative traces and amplitudes of EPSCs recorded in DMH neurons following light stimulation (473 nm, 5 ms) of MPOBDNF afferents. w Representative traces and amplitudes of EPSCs recorded in TrkB-knockdown DMH neurons after HA training in MPOBDNF-ChR2 mice. x Summary of the role of BDNF in HA. All data are shown as mean ± SEM and analyzed by t-test. *P < 0.05, **P < 0.01, ***P < 0.001 vs corresponding control group; ns, not significant.
Building on previous findings that BDNF neurons in the preoptic area (POA) are crucial for heat defense and that acute heat exposure significantly increases POA BDNF expression,6,7 we hypothesized that BDNF plays a role in HA. In line with this, HA mice showed elevated Bdnf mRNA expression after subsequent 38 °C exposure (Fig. 1d). Considering the importance of intrinsic thermosensitivity in heat defense8,9 and its proposed role in HA,10,11 we first recorded the basal firing rates at 36 °C and found that HA notably increased the firing rates of MPOBDNF neurons. We further found that the thermosensitivity of MPOBDNF neurons increased significantly after HA, resulting in a rise in the proportion of warm-sensitive neurons (WSNs, defined as neurons with thermosensitivity > 0.8 imp/s/°C). The proportion increased from 22.7% to 43.3% (Fig. 1e), exceeding the overall increase in MPO neurons (from 27.3% to 36.7%; Supplementary information, Fig. S4a). Taken together, these results highlight that HA increases MPO BDNF expression, and intrinsic thermosensitivity of MPOBDNF neurons.
To determine the roles of MPOBDNF neurons in HA, we inhibited them using tetanus neurotoxin (TeNT) (Fig. 1f). Consistent with their reported role in heat defense,7 this inhibition impaired basal heat defense function, leading to a rise in Tcore at 38 °C during the first day of HA training (Supplementary information, Fig. S4b, c). Notably, this inhibition nearly abolished the HA effect on Tcore tolerance, with Tcore rapidly increasing to 41.5 °C during the HTT (Fig. 1g). Concurrently, the anxiety-reducing effect of HA was also negated (Fig. 1h; Supplementary information, Fig. S4d). Thus, MPOBDNF neurons are required for the HA effects on Tcore and anxiety.
To ascertain whether MPOBDNF neural activation could induce an HA effect without HA training, we employed optogenetic activation using ChR2 (Fig. 1i). Intriguingly, optostimulation at 1 Hz over three consecutive days elicited a pronounced HA effect, as evidenced by a 2-fold increase in the time taken to reach 41.5 °C in the HTT compared with GFP controls (Fig. 1j). To identify downstream targets of MPOBDNF neurons, we considered the DMH and rRPa (Fig. 1k), as suggested in the thermoregulatory circuitry.12 We then employed a projection-specific strategy to block synaptic transmission (Fig. 1l). As expected, blocking either DMH- or rRPa-projecting MPOBDNF neurons had a small but significant effect on basal heat defense function during 38 °C exposures (Supplementary information, Fig. S4e, f). Notably, blocking either of these two pathways nearly abolished the HA effect on Tcore tolerance and anxiety alleviation during heat exposures (Fig. 1m, n; Supplementary information, Fig. S4g, h). These data collectively demonstrate that the MPOBDNF → DMH/rRPa neurocircuitry is essential for the HA effect.
To determine the role of BDNF in HA, we knocked down its expression by injecting Lenti-Bdnf-shRNA virus into the MPO of C57 mice (Fig. 1o). BDNF knockdown in the MPO did not alter the basal heat defense function (Supplementary information, Fig. S4i–k). Surprisingly, BDNF knockdown had no effect on Tcore tolerance during HTT (Fig. 1p). In contrast, the knockdown compromised the anxiolytic effects of HA, as indicated by reduced open arm exploration in the EPM (Fig. 1q) and reduced central exploration in the OFT (Supplementary information, Fig. S4l). Therefore, BDNF in the MPO is essential for the anxiolytic effects of HA.
To determine whether the BDNF receptor TrkB is involved in HA, we knocked it down in various regions axonally projected by MPOBDNF neurons, including the DMH, rRPa, paraventricular thalamus (PVT), periaqueductal gray (PAG), and mamillary peduncle (mp) (Fig. 1r; Supplementary information, Fig. S5a, b). Similar to BDNF knockdown in the MPO, TrkB knockdown in the DMH did not affect basal heat defense when exposed to 38 °C (Supplementary information, Fig. S5c–e). Additionally, it had no effect on Tcore tolerance during HTT (Fig. 1s). However, it significantly reduced the anxiolytic effects of HA, as evidenced by decreased open arm exploration in the EPM (Fig. 1t) and reduced central exploration in the OFT (Supplementary information, Fig. S5f). Beyond the DMH, knockdown in none of the other regions — rRPa, PVT, PAG, or mp — affected HA’s impact on Tcore or anxiety (Supplementary information, Fig. S5g–j). Further, the simultaneous knockdown of TrkB in the DMH and rRPa did not affect Tcore tolerance (Supplementary information, Fig. S5k). In conclusion, TrkB in the DMH is selectively required for the anxiolytic effects of HA.
Synaptic plasticity is postulated as a pivotal mechanism underlying HA.10 Given that BDNF is known to modulate synaptic plasticity,13 we suspected that BDNF-dependent synaptic remodeling within the BDNF circuitry could be instrumental for HA. Previous studies documented that MPOBDNF neurons consist of mixed glutamatergic and GABAergic subpopulations, with proportions of 60% and 33%, respectively.6,7 We confirmed both connections by recording excitatory and inhibitory postsynaptic currents (EPSCs/IPSCs) in DMH neurons, evoked by optogenetic stimulation of MPOBDNF terminals in the DMH (Fig. 1u; Supplementary information, Fig. S6a). Our data showed that 64%–67% of neurons had light-evoked EPSCs, 62% had light-evoked IPSCs, and 26%–29% exhibited both EPSCs and IPSCs in either HA or control groups (Supplementary information, Fig. S6b). However, the amplitude of light-evoked EPSCs increased (Fig. 1v), while that of IPSCs decreased in HA mice (Supplementary information, Fig. S6c), suggesting potentiation of excitatory and depression of inhibitory connections in the MPOBDNF → DMH pathway.
To discern whether this potentiation was pre- or postsynaptic, we analyzed the paired-pulse ratio (PPR) of two light-evoked EPSCs within a 50-ms interval and characterized properties of miniature EPSCs/IPSCs (mEPSCs/mIPSCs). There was no difference in PPR or mEPSC frequency between groups, indicating no change in presynaptic glutamate release probability (Supplementary information, Fig. S6d, e). However, there was a marked increase in the amplitude of mEPSCs (Supplementary information, Fig. S6e), pointing to an increase in postsynaptic ion conductance. In contrast, both the amplitude and frequency of mIPSCs remained unchanged (Supplementary information, Fig. S6f). Further quantal EPSC recordings with strontium14 revealed that HA mice exhibited increased amplitude but unchanged frequency of quantal EPSCs compared to controls (Supplementary information, Fig. S6g). Knocking down TrkB in DMH neurons abolished HA-mediated EPSC potentiation (Fig. 1w; Supplementary information, Fig. S6h), but did not affect IPSC amplitude (Supplementary information, Fig. S6i). Taken together, we conclude that the MPOBDNF → DMH pathway undergoes TrkB-dependent potentiation of excitatory transmissions primarily via increased postsynaptic ion conductance.
In summary, we establish a simple and easily replicable HA protocol in mice and explore HA’s profound effects on thermoregulation and anxiety alleviation. These effects are primarily mediated by the MPOBDNF → DMH/rRPa thermoregulatory pathways (Fig. 1x). Within these pathways, HA enhances BDNF expression in the MPO and increases neuronal intrinsic thermosensitivity. These BDNF molecules interact with TrkB in the DMH, potentiating excitatory synaptic connectivity and alleviating anxiety. Additionally, it is worth noting that BDNF levels in human serum also increase following heat exposure,15 suggesting a potentially conserved mechanism between mice and humans. Thus, our research not only deepens our understanding of physiological adaptations to heat but also informs strategies to improve thermal performance.
留言 (0)