
記住我
During learning experience, a subset of astrocytes in the hippocampus is activated and becomes both necessary and sufficient for subsequent memory recall. Williamson et al. discovered that these learning-associated astrocytes play an important role in memory by modulating engram neurons’ synaptic activity and facilitating the reactivation of engram neuron ensembles during memory retrieval.
Memory allows us to store and retrieve previously acquired information. Many results from rodent studies show that memories are stored in sparse populations of neurons that are active during learning and are reactivated during memory recall, termed engram ensembles.1 Because neuronal activity at the time of learning is a hallmark of neurons in an engram ensemble, the use of promoters for activity-dependent immediate early genes (IEGs) such as cFos allows the visualization and manipulation of engram neurons across brain regions and different types of memory. However, these studies largely focused on the role of excitatory neurons in engram ensembles.2
Recent studies suggest a role of non-neuronal cells, including astrocytes, in memory.3,4 Traditionally, astrocytes were thought to function as neuronal “supporters” that encapsulate neurons to regulate ion concentration and neurotransmitters in the extracellular space.5 Interestingly, astrocytes express similar receptors and voltage-gated ion channels to neurons. Once activated, astrocytes undergo an increase in the intracellular calcium level which leads to the secretion of signals that enhance neuronal synaptic function.5 The first hint that astrocytes play a role in memory was the finding that activation of astrocytes in the dorsal hippocampal CA1 region of mice enhanced memory.6 This group also showed that inhibiting astrocytes in this region during training impaired memory retrieval when mice were tested at remote times.7 These findings inspired some intriguing questions: Do astrocytes act in synergy with engram neurons in the hippocampus to mediate memory formation and recall? And if so, are there specific populations of astrocytes that are critical to one particular memory similar to engram neurons?
A new study from Williamson and colleagues discovered that only a subset of astrocytes in the dentate gyrus and CA1 regions of the dorsal hippocampus express cFos after learning, naming these learning-associated astrocytes (LAAs).8 But is this experience-dependent and sparse activation of astrocytes important for that memory? To test this, the authors knocked out cFos from hippocampal astrocytes by microinjecting a GFAP-Cre virus (to target astrocytes) into the hippocampus of mice that have a floxed Fos allele. Mice were trained in contextual threat conditioning in which a context is paired with an aversive footshock and tested by replacing mice in the training context without footshock. Knocking out cFos from hippocampal astrocytes prevented the induction of cFos expression normally observed during learning and impaired memory retrieval. Moreover, hippocampal slices from these mice showed a decrease in long-term synaptic potentiation. These findings suggest that much like engram neurons, a subpopulation of astrocytes is active during learning and become essential for subsequent memory recall (Fig. 1).
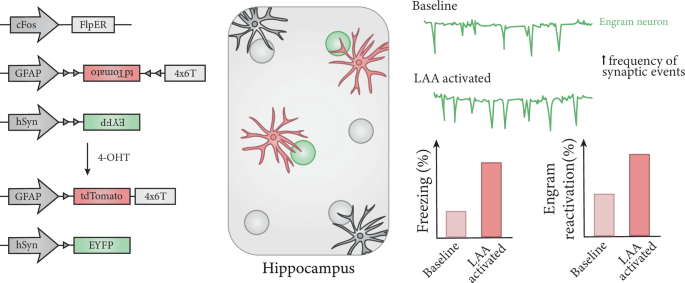
Fig. 1: Collaborative role of LAAs and engram neurons in memory.
Viruses with activity (cFos promoter)-dependent flippase (FlpER) alongside astrocyte- and neuron-specific promoters (GFAP and hSyn, respectively) were used to tag LAAs (red, star) and engram neurons (green, circle) in the hippocampus. 4×6T microRNA targeting cassette improves astrocyte specificity, while 4-hydroxytamoxifen (4-OHT) is used to induce active cell labeling. LAAs preferentially located near engram neurons. Artificial activation of LAAs during learning facilitates synaptic transmission in engram neurons, induces memory retrieval and increases the chance of engram reactivation during memory test.
Williamson and colleagues next examined whether LAAs interact with engram neurons during memory acquisition and retrieval using a clever viral strategy to label both astrocytes and neurons active during threat training. One virus expressed tamoxifen-inducible flippase downstream of cFos promoter to label both neurons and astrocytes active during learning. To differentiate between these cell types, they used two additional viruses with either astrocyte- or neuron-specific promoter containing flippase-dependent fluorescent reporters in different colors. Therefore, administration of 4-hydroxytamoxifen during contextual threat conditioning would tag both engram neurons and LAAs. This unique combination of methods allowed the authors to show that engram neurons preferentially colocalize with LAAs rather than with non-LAAs. Moreover, in slice physiology experiments, they found that artificially activating LAAs that were labeled during threat conditioning enhanced synaptic transmission in engram, but not non-engram, neurons. This suggests that the activation of astrocytes induces a long-term potentiation-like phenomenon in engram neurons. Together, these results suggest both structural and functional communication between LAAs and engram neurons that are important for memory.
Memory retrieval is induced by an external sensory cue reactivating the neuronal engram ensemble. Studies show that artificial activation of the neuronal engram ensemble can induce memory retrieval even in the absence of external sensory retrieval cues.9 The authors examined whether artificial excitation of LAAs is also sufficient to induce memory retrieval in the absence of sensory retrieval cues. First, they expressed the excitatory designer receptor exclusively activated by designer drugs (DREADD) in LAAs by administering tamoxifen during threat conditioning as above. To test memory, the DREADD agonist was delivered and mice were placed in a novel context (in which they had not previously experienced a footshock). Mice showed freezing behavior indicative of artificial retrieval of a threat memory. Importantly, no freezing was observed in mice that expressed the DREADD receptor in a random population of astrocytes. Furthermore, this effect was durable and was observed a month after training. Follow-up experiments showed that artificially activating LAAs induced activity in the neuronal engram ensemble, suggesting that astrocytes help reactivate the neuronal engram ensemble to induce memory retrieval.
The paper by Williamson et al. opens up the intriguing possibility that astrocytes are critical components of an engram ensemble that supports a specific memory. It provides an essential bridge between the roles of neurons and glial cells in engram ensembles by showing that engram neurons and their associated LAAs work hand-in-hand to support contextual threat memory formation and retrieval. The findings presented in this study also prompt some outstanding questions. First, do these LAAs store precise information or are they once again relegated to the role of supporting neuronal engram ensembles? Second, is this role of astrocytes in engram ensembles specific to the hippocampus and contextual memories or does it generalize to other brain regions and other types of memory? Third, what is the nature of the interaction between astrocytes and neuronal engram ensembles? With these provocative findings, Williamson and colleagues provide new insights into the role of non-neurons in engram ensembles supporting memories.
留言 (0)