
記住我
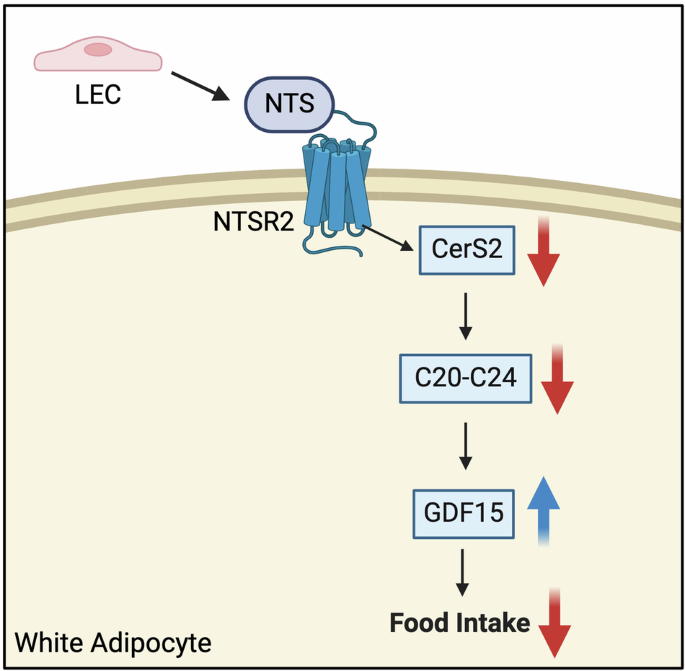
In a recent work published in Nature, Wang et al. reveal how protein kinase A and calcineurin dynamically regulate the daily duration of sleep and wake periods at excitatory post-synapses in the brain. This study uncovers key molecular mechanisms offering new insights into the enzymatic control of sleep homeostasis.
We all need sleep, but what exactly drives our need for sleep? We do know that the need to sleep is most profoundly determined by the time spent awake.1 The longer we are awake the more urgent the need to sleep is, with this experience reflecting the homeostatic regulation of sleep. The regulation of sleep is additionally shaped by the circadian rhythm — sleep pressure is higher in the evening when we are used to going to bed, than in the morning.
Wang et al.2 have now provided novel insights into the molecular factors underlying these basic principles of sleep regulation. Developing over 40 knockout mouse lines, they identified key protein kinases and phosphatases involved in the brain’s control of sleep and wakefulness, pinpointing protein kinase A (PKA) and calcineurin subunits as critical signals in this process. The authors used adeno-associated virus (AAV)-mediated stimulation of PKA and calcineurin in mice, demonstrating that PKA acts to promote wakefulness, whereas the phosphatase calcineurin serves to promote sleep. Remarkably, the manipulations of PKA and calcineurin resulted in an up to 4-fold increase or decrease in sleep duration, respectively. Moreover, the authors could allocate the effects of PKA and calcineurin to the post-synaptic level of excitatory glutamatergic synapses, where the two enzymes appear to compete.
The findings by Wang et al. are remarkably well in line with the Synaptic Homeostasis Hyposthesis (SHY), assuming that sleep serves the global re-normalization of synapses.3 Enhanced encoding of information during the wake period leads to an increase in synaptic potentiation and connectivity in the brain’s glutamatergic networks, and sleep, in turn, reduces synaptic strength, thus helping restore a balance in network excitability. This global synaptic re-normalization is associated with the slow wave sleep (SWS) stage of sleep, rather than a consequence of rapid eye movement (REM) sleep.4 Fittingly, in Wang et al.’s mouse lines, changes in PKA and calcineurin activities were not only reflected in differences in sleep duration but also in delta power, i.e., the amplitude of the 0.5–4 Hz slow waves that hallmark the brain’s electrical activity during SWS.
The experiments by Wang et al. unmasking the neuronal processes underlying sleep-wake regulation at an unprecedented molecular level, inevitably prompt several basic questions. One concerns the contribution of circadian factors, as many of the observed effects depended on the time of day. However, previous work has shown that sleep deprivation abolishes nearly all phosphorylation cycles in synaptoneurosomes,5 suggesting that the post-synaptic enzymatic processes indeed reflect the homeostatic, rather than circadian, regulation of sleep.
Against the backdrop of SHY, a more pressing question is how the observed post-synaptic enzymatic processes link to the electrical slow wave rhythm during SWS that has likewise been considered to causally contribute to the re-normalization of excitatory synapses during sleep.3 Does post-synaptic calcineurin activity acutely support the emergence of electrical slow waves or, conversely, does the emergence of slow wave activity acutely change post-synaptic activity of these enzymes? Beyond acute interactions, a loop-like scenario is conceivable, such that PKA activity promotes wakefulness and arousal, at the same time supports synaptic long-term potentiation (LTP) and, thus, a global upscaling of excitatory synapses during wakefulness. The resultant net increase in synaptic connectivity, in turn, enables the occurrence of slow waves during subsequent sleep, accompanied by an increased post-synaptic calcineurin activity. Calcineurin, indeed, fosters long-term depression (LTD),6 and may thus contribute to global synaptic downscaling processes during sleep (Fig. 1). Interestingly, the slow waves of SWS, in addition to their involvement in global synaptic re-normalization, help consolidate newly formed memories, i.e., a process associated with shaping synaptic connections within specific circuits.7 And the competitive regulation of PKA and calcineurin during sleep may likewise enable such dual function of globally re-normalizing synaptic networks and, in parallel, stabilizing synaptic connectivity in circuits underlying specific memories.6
Fig. 1: Two key questions arising from Wang et al.2
Left: Does predominant post-synaptic calcineurin activity drive slow wave activity (SWA) during SWS, or vice versa? Right: Does the effect of post-synaptic PKA/calcineurin competition on sleep regulation reflect a global feedback effect of this competition on hypothalamic sleep regulatory circuits, or a local effect of the enzyme competition on hypothalamic sleep circuits (e.g., in the VLPO)?.
How do the changes in post-synaptic enzyme activity as mechanism mediating sleep duration, connect to what is known about the local mechanisms of sleep-wake regulation that particularly rely on specific cell circuits in the ventrolateral preoptic nucleus (VLPO) and the lateral hypothalamus?8 Wang et al.’s genetic approach implicates a wholistic concept of sleep–wake generation in which enzyme activity at basically every excitatory post-synapse in the brain potentially feeds into the regulation of sleep. This apparently contrasts with the idea that sleep emerges from quite localized circuits in the hypothalamus. Reconciling these views, one might assume that not every post-synapse in the brain is equally relevant but, that the influence of PKA/calcineurin interactions on sleep duration is conveyed specifically by synaptic circuits that are located in the hypothalamic sleep regulating centers (Fig. 1). This possibility seems in line with evidence suggesting that hypothalamic circuits directly involved in sleep–wake control undergo AMPA receptor downregulation together with reduced phosphorylation of S845 on the GluR1 subunit during SWS.4 Wake-associated synaptic upscaling in hypothalamic networks, on the other hand, might directly promote sleep by increasing activity levels of sleep-promoting cell populations. Indeed, phosphorylation of S845 on GluR1 as a substrate of PKA enables the incorporation of AMPA receptors into post-synapses everywhere in the brain.9 Accordingly, enhanced PKA activity is expected to increase activity levels at glutamatergic excitatory synapses also in the sleep centers of the hypothalamus. Alternatively, functions of the hypothalamic sleep regulating circuits may essentially rely on input deriving from PKA-induced changes in synaptic plasticity in extrahypothalamic, e.g., thalamo-cortical regions.10
Ultimately, the experiments by Wang et al. prompt the questions of what is cause and effect in sleep–wake regulation, at two different levels: Do changes in post-synaptic enzyme activity cause changes in electrical slow wave activity, or vice versa? And, do changes in post-synaptic enzyme activity cause global or local changes in hypothalamic sleep regulatory centers? Loop-like interactions between the processes are conceivable as well. Despite a fascinating endeavor, addressing these questions will be challenging since it requires acute and local manipulations of post-synaptic enzyme activity in conjunction with sleep and slow waves.
留言 (0)