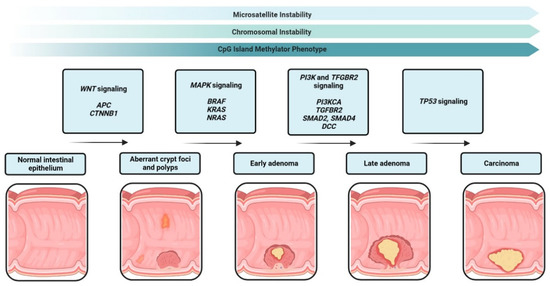
CRCs arise from precursor polyps, mostly from the two most common polyps, pre-cancerous conventional adenomas and sessile serrated polyps [
80]. One hallmark of tumorigenesis is the accumulation of mutations or epigenetic alterations. These mutations and alterations lead to the activation of oncogenes or the inactivation of tumor suppressor genes. However, this is not enough for malignant cell transformation, as some studies have shown that phenotypically normal cells can carry several somatic mutations without developing into cancer cells [
81,
82,
83,
84]. For the progression of normal cells into malignant cells with uncontrolled infinitive proliferation capacity, cells of the microenvironment have to support and promote tumorigenesis by pro-inflammatory cytokines. As mentioned in the previous paragraph, inflammatory cytokines support successful tumor development, transformation and progression of mutated cells. In addition, as also mentioned in the previous paragraph, most pro-inflammatory cytokines are necessary for gut homeostasis and wound healing. Examples for these dichotomous functions are IL-1α and IL-1β. As alarmins that sense DAMPs and PAMPs, IL-1α and IL-β activate a cascade of cytokines. This cascade plays a major role in the activation and orchestration of innate and adaptive immunity. Many of the intrinsic (oncogene-driven) and the extrinsic (TME-driven) tumor-promoting effects are mediated by IL-1 [
85]. IL-1β mediates cell proliferation, differentiation and apoptosis; stimulates the expression of TNF, IL-6, IL-8, and IL-17A; and increases Wnt/β-catenin signaling, promoting intestinal tumorigenesis [
37]. After a localized inflammatory process with the secretion of IL-1 cytokines from, e.g., macrophages and neutrophils, CRC tumor cells can create a positive feedback loop, promoting the expression of more pro-inflammatory cytokines that stimulate cancer-cell proliferation and drug resistance [
36]. The regulation of cytokines is quite complex as cytokines are not only regulated by transcriptional and post-transcriptional mechanisms but also by the availability of cytokine receptors, which are also regulated transcriptionally and post-transcriptionally.
Table 1.
Cytokines in colorectal cancer.
Cytokines that Promote CRC Cytokine FamilyCytokineReceptorFunctional effect in CRCReferenceIL-1 FamilyIL-1αInterleukin 1 receptor type 1 (IL-1R1)-IL-1R3Promotes inflammatory carcinogenesis[
86] IL-1βIL-1R1-IL-1R3IL-1R2-IL-1R3Promotes the proliferation of colon cancer cells, promotes inflammation-induced carcinogenesis[
86,
87]IL-2 (common γ-chain) FamilyIL-4IL-4RPromotes Th2-type inflammation and Th9 polarization[
88,
89]IL-6 FamilyIL-6IL-6Rα-gp130Promotes carcinogenesis via upregulation of proliferation, mitosis, migration and angiogenesis[
90,
91,
92] IL-11IL-11Rα-gp130Promotes inflammation-induced carcinogenesis, facilitates the proliferation of colon cancer cells[
93,
94,
95] IL-31IL-31Rα-OSMRbPromotes Th2 cell polarization, evidently tumorigenic[
96]IL-8 FamilyIL-8CXCR1, CXCR2Promotes colon cancer cell proliferation, attracts neutrophils, mediates a suppressive environment and chemoresistance[
97,
98,
99,
100]IL-10 FamilyIL-19IL20Rα-IL20RβEvidently tumorigenic[
101] IL-20IL-20Rα-IL-20RβPromotes carcinoma outgrowth, induces PD-1[
102] IL-22IL-22Rα-IL-10RβPromotes progression of carcinomas, stemness and proliferation[
103,
104,
105] IL-26IL-20Rα-IL-10RβPromotes TH17 polarization, only expressed in humans, not in mice[
102]IL-12 FamilyIL-23IL-23R-IL-12RβPromotes pro-inflammatory cytokine secretion[
106,
107] IL-35IL-12Rβ-gp130
IL-12Rβ-IL-12RβPromotes Treg-mediated suppression of T cell responses and exhaustion of T cells[
108]IL-17 FamilyIL-17AIL-17RA-IL-17RCPromotes cell cycle progression of colon cancer cells and immunosuppression[
109,
110,
111]Other cytokines IL-13IL-13Rα-IL-4RαPromotes Th2 cell polarization[
112] IL-16CD4Evidently pro-tumoral[
113] IL-34CSF1RPromotes cancer progression and immune suppression and therapeutic resistance[
114,
115,
116] TNFTNFR1, TNFR2promotes inflammation[
117,
118,
119]ChemokinesCXCL-1CXCR1, CXCR2Recruitment of tumor-associated macrophages (TAM) and TANs[
120,
121]CXC chemokinesCXCL-2CXCR2Recruitment of TAMs and TANs[
122] CXCL-8CXCR1, CXCR2Recruitment of TANs, TAMs and cancer cells to the tumor site; promotion of angiogenesis and tumor stemness[
123,
124,
125] CXCL-12CXCR4, CXCR7Recruitment of TAMs at the invasive front, upregulation of IL-10[
126,
127] CXCL-16CXCR6Induces the polarization of macrophages toward a pro-tumoral M2 phenotype[
128]CC chemokinesCCL-2CCR2Recruitment of TAMs at the invasive front, recruitment of myeloid-derived suppressor cells (MDSC) into the tumor[
129,
130] CCL-3CCR1, CCR4Recruitment of MDSC into the tumor, promotes proliferation of colon cancer cells[
131] CCL-4CCR1, CCR3Recruitment of MDSC and TAMs into the tumor[
132] CCL-11CCR8Promotes tumor angiogenesis, inhibits apoptosis of endothelial cells[
133] CCL-16CCR1, CCR2, CCR3, CCR5, CCR8Promotes tumor angiogenesis, inhibits apoptosis of endothelial cells[
134] CCL-17CCR4Recruitment of Tregs and Th2 lymphocytes[
135] CCL-20CCR6Recruitment of Tregs and Th2 lymphocytes[
136,
137]
Cytokines with a more CRC-suppressive phenotype Cytokine FamilyCytokineReceptorFunctional effect in CRCReferenceIL-1 FamilyIL-18IL-5R5-IL-1R7Activates lymphocytes to produce IFN-g, restricts ThH17 differentiation[
138,
139] IL-36IL-1R6-IL1R3Conservation of tertiary lymphoid structures[
34,
140,
141] IL-37IL-1R8-IL-1R5Inhibits β-catenin[
141,
142]IL-2 (common γ-chain) FamilyIL-2IL-2RT and NK cell activation factor[
34,
143] IL-7IL-7RαPromotes the proliferation of T cells and NK cells[
144,
145] IL-9IL-9RaPromotes the proliferation of CD8+ T cells[
146,
147] IL-15IL-15-IL-15RαPromotes the proliferation and activation of NK cells and CD8+ T cells[
148,
149,
150] IL-21IL-21RPromotes the proliferation and activation of NK cells and CD8+ T cells[
151,
152,
153]IL-10 FamilyIL-24IL-20Rα-IL-20Rβ,
IL-22Rα-IL-20RβInduces apoptosis and autophagy of cancer cells[
154]IL-12 FamilyIL-12IL-12RβPromotes T cell survival and proliferation and the proliferation of NK cells, enhances cytotoxic function[
155,
156]IL-17 FamilyIL-17FIL-17RA-IL-17RCInhibition of tumor angiogenesis, enhancing immune cell recruitment[
34,
157]Other cytokines Interferon FamilyIFN-γIFNGRMediates anti-proliferative, anti-angiogenic and pro-apoptotic effects[
44,
158,
159] Type I interferonsInterferon alpha and beta receptor subunit 1 (IFNAR1)-IFNRA2Recruitment of CD8+ lymphocytes[
160]TGF-βTGF-βTGFBR1-TGFBR2Inhibits cancer cell proliferation, regulates immune cell differentiation[
161,
162]Chemokines CXC chemokinesCXCL-9CXCR3Promotes the infiltration of NK cells, CD8+ and CD4+ T cells into the tumor, suppression of angiogenesis[
163,
164] CXCL-10CXCR3Promotes the infiltration of NK cells, CD8+ and CD4+ T cells into the tumor, suppression of angiogenesis[
160,
163,
165] CXCL-11CXCR3Promotes the infiltration of NK cells, CD8+ and CD4+ T cells into the tumor, suppression of angiogenesis[
166]
Cytokines with a mixed phenotype Cytokine FamilyCytokineReceptorFunctional effect in CRCReferenceIL-1 FamilyIL-33IL-1R4Alters TME, promotes angiogenesis, enhances colon cancer stemness but induces anti-tumoral IFN-γ responses[
167,
168,
169,
170]IL-2 (common γ-chain) FamilyIL-9IL-9RInhibits tumor growth by enhancing immune responses and promotes tumor growth by suppressing immune responses[
112,
171]IL-10 FamilyIL-10IL-10Rα-IL-10RβPromotes cytotoxicity but inhibits anti-tumor responses[
172]Chemokines CXC chemokinesCXCL-4 Suppresses the activity of CD8+ T cells and inhibits tumor angiogenesis and endothelial cell proliferation[
173] CCL-5CCR1, CCR3, CCR4, CCR5Recruitment of MDSC and fibroblasts into the tumor but also CD8+ T cells[
160,
174,
175] 2.2.1. Cytokines in the Early Stages of CRCDisruption of the epithelial barrier by bacterial infection, microbial metabolites, obesity or epithelial damage results in the production and release of several pro-inflammatory cytokines. However, even the alteration in gut microbiota (
Figure 3) can promote tumorigenesis as the microbiome and the intestinal epithelial cells interact in a complex network to maintain homeostasis [
176]. The gut microbiome of a healthy individual is characterized by a high species diversity of, e.g., Bacteroidetes, Firmicutes and Actinobacteria [
177,
178]. Colorectal cancer is associated with a dysbiosis of this diversity with an overrepresentation of specific bacteria such as the anaerobic bacterium Fusobacterium nucleatum [
179], Streptococcus gallolyticus, Bacteroides fragilis, Escherichia coli or Enterococcus faecalis [
180]. Gut bacteria can influence tumor growth or inhibition of CRC proliferation through direct cell interaction and through microbial-derived metabolites [
181]. The most well studied metabolites are the short-chain fatty acids (SCFAs). SCFA metabolism by epithelial cells maintains the hypoxic environment of the colon that most bacteria require. In the absence of SCFA, epithelial cells switch to anaerobic respiration, release oxygen and nitrates into the colon lumen and fuel the expansion of pathogens such as Escherichia coli [
182]. Escherichia coli is a Gram-negative bacterium with a membrane that consists of lipopolysaccharides (LPS). LPS bind to the Toll-like receptor (TLR) 4 on many immune and some epithelial cells and induce the secretion of pro-inflammatory cytokines. TLRs are a family of pattern-recognition receptors mostly expressed on the surface of cells of the innate immunity such as granulocytes. SCFAs prevent tumorigenesis by inhibiting LPS-induced expression of pro-inflammatory cytokines and induce colon adenoma and carcinoma cell apoptosis [
183]. SCFAs decrease the two important CRC signaling pathways nuclear factor kappa-B (NF-κB) and Wnt, facilitating anti-tumorigenic effects, and reduce pro-inflammatory cytokines such as TNF, IL-6 or CCL-3 [
181]. Some pathogenic bacteria are able to invade the epithelium directly by increasing the intestinal permeability. This is achieved by binding to the tight junction proteins such as E-cadherin, occludin or claudin1. Fusobacterium nucleatum can bind to E-cadherin and invade the epithelial cell, thus, promoting colon cancer [
184]. There are some more microbial modulators of tumor inflammation such as microbial toxins that activate immune responses and inflammation. For example, the Bacteroides fragilis toxin (BFT) and the colibactin from Eschericchia coli induce DNA damage in epithelial cells (referred as genotoxins) and, subsequently, a strong TH17 response [
185,
186]. Conversely, bacterial adhesins such as FadA from Fusobacterium nucleatum or AFA-1 from Eschericchia coli can bind E-Cadherins on the surface of epithelial cells and activate β-catenin signaling and, subsequently, the release of IL-6 and TNF [
187].Disrupted intestinal barriers at the site of a small colorectal tumor induce the activation of innate immune cells and the increased expression of pro-inflammatory cytokines. One of the major cytokines that fuels CRC progression is TNF. TNF is one of the earliest and most important pro-inflammatory cytokines that activates other pro-inflammatory cytokines via the NF-κB signaling pathway. TNF has also contradictory effects in cancer progression due to its two receptors. TNFR1 has a cytoplasmic death domain so that binding of TNF to the TNFR1 can lead to apoptosis [
188]. In CRC, the apoptosis-inducing properties of TNF are less pronounced compared with the pro-inflammatory properties that induce the production of several inflammatory cytokines, enhanced oncogene activation, tumor cell invasion and migration and creation of a tumor-supportive TME [
172]. Concomitant with or shortly after the secretion of TNF, the cytokines IL-1 and IL-6 are produced. As described in the former paragraphs, the alarmins IL-1α and IL-1β mediate cell proliferation, differentiation and apoptosis, stimulate the expression of TNF, IL-6, IL-8, and IL-17A, and increase the Wnt/β-catenin signaling [
37].The cytokine IL-6 is involved not only in cancer inflammation but also in hematopoiesis, bone metabolism and embryonic development [
92]. The IL-6R is composed of the binding receptor chain IL-6Ra and the signal transducer chain glycoprotein 130 (gp130). As IL-6 is a target of the transcription factor NF-κB, the activation of NF-κB by, e.g., TNF, simultaneously with the activation of signal transducer and activator of transcription 3 (STAT3) in non-immune cells such as epithelial cancer cells, triggers a positive feedback loop by the IL-6-signal transducer and activator of transcription 3 (STAT3) axis [
68]. The resulting excessive activity of IL-6 leads to the overexpression of the proto-oncogene c-Myc, multiple pro-inflammatory cytokines and, consequently, tumor growth, tumor progression and drug resistance. At the same time, IL-6 suppresses anti-tumorigenic immune responses [
189]. In the early stages of CRC, chemokines shape the composition of the TME. The chemokine IL-8 attracts neutrophils and can be induced by TNF. TH17 cells express elevated levels of the C-X-C chemokine receptor type 4 (CXCR4), the receptor of CXCL-12 and CC chemokine receptor 6 (CCR6), the receptor of CCL-20. The chemokines that attract monocytes are CCL-2 and CXCL-5 [
43]. Thus, at the early phase of colorectal cancer, cytokines change the balance as the TME develops to a more tumor promoting or tumor suppressing immune microenvironment. 2.2.2. Cytokines in the Late Stages of CRCAs the tumor progresses, more immune cells enter the tumor, attracted by chemokines. TNF and IL-1 synthesized by leukocytes renders endothelial cells into pro-inflammatory endothelial cells that secrete a large amount of chemokines, such as IL-8 and CXCL-2, to further enhance the infiltration of neutrophils. As neutrophils are the first responders against extracellular pathogens, neutrophils were originally considered to be defensive against colorectal cancer [
190]. However, neutrophils in CRC can promote the growth of cancer cells by regulating the innate and adaptive immune system and inducing angiogenesis [
191]. In the TME, neutrophils can differentiate into tumor-associated neutrophils (TANs) with a N1 or N2 phenotype (see
Figure 4 for an overview of cells and cytokines of the TME). N1 TANs have a more anti-tumorigenic phenotype, whereas N2 TANs have a more tumor-promoting phenotype. TGF-β polarizes toward a N2 phenotype that produces several chemokines such as CCL-2, CCL-5 and CXCL-4 [
191]. IFN-β polarizes toward an N1 phenotype [
192].Fibroblasts play a key role in wound healing as they are the major producers of the extracellular matrix. Although cancer-associated fibroblasts (CAFs) are a central component of the TME of nearly every solid tumor, they comprise a heterogeneous population of cells with no precise fibroblast-specific markers. Cells that are negative for epithelial, endothelial and leukocyte markers with an elongated morphology and lacking the mutations of cancer cells are considered to be CAFs [
193]. In the TME, various inflammatory cytokines such as IL-1, TNF or IL-6 can activate fibroblasts to become CAFs. CAFs secrete numerous cytokines and chemokines, such as TGF-β, IL-6, IL-1β, IL-4, and CXCL-12 (SDF-1), which influence CD8+ T cells, Tregs and macrophages to act as immuno-suppressive or immuno-promoting cells. In CRC, CAFs have more immuno-suppressive effects and consequently promote cancer cell proliferation and therapy resistance [
194].Tumor-associated macrophages (TAMs) also play a major role in the TME of CRC. Macrophages eliminate invading microbes by phagocytosis, in homeostasis resident macrophages in the colon secrete high amounts of IL-10 and are, therefore, anti-inflammatory [
195]. Under inflammatory conditions, inflammatory CX3CR1-positive macrophages enter the colon and secrete the pro-inflammatory cytokines TNF and IL-6 [
196]. Similar to neutrophils, macrophages can differentiate into two different phenotypes. The classically activated M1 phenotype has the capability to promote T helper 1 (TH1) responses and kill tumor cells. M1 differentiation is induced by bacterial components, IFN-γ and TNF. In contrast, alternatively activated M2 macrophages display tumor-promoting activity and immuno-suppressive functions. M2 differentiation is induced by IL-6, IL-10 or IL-13. M2 macrophages are typically divided into four subtypes, which are differently stimulated: the M2a, induced by IL-4 and IL-23; the M2b subtype, stimulated by IL-1; the M2c subtype, induced by IL-10 and the M2d macrophage, induced by IL-6. Through the expression of the two immuno-suppressive cytokines IL-10 and TGF-β, M2 macrophages directly induce an immunosuppressive microenvironment [
197,
198]. Through the secretion of IL-6, M2 macrophages mediate the expression of IL-10 by colorectal cancer cells and indirectly induce a microenvironment that suppresses T cell activity [
199]. The partition of macrophages in the two subtypes M1 and M2 is a simplified view; in reality, TAMs exist in various states between the two phenotypes.Macrophages and neutrophils belong to the group of tumor-infiltrating myeloid cells (TIMs), a heterogeneous population of cells characterized by diversity and plasticity. One member of this myeloid cell group is myeloid-derived suppressor cells (MDSCs). MDSCs are able to most effectively suppress T cell activities by secreting high amounts of IL-10 and TGF-β [
200]. MDSCs are very plastic as they can differentiate into TAMs or tumor-associated DCs, depending on the signals in the TME [
201].
T cells are the most abundant in the TME and can be divided into CD8+ cytotoxic T lymphocytes (CTLs), CD4+ T helper cells, regulatory T cells and NKT cells.
CD8+ CTLs are effector cells of the adaptive immune system and play a pivotal role in anti-tumorigenic immune responses by direct killing of malignant cells. CTLs in the TME of CRC produce high amounts of IL-2, IL-12 and IFN-γ that activate the killing efficiency of NK cells and CTLs [
202] and enhance the expression of CXCL-9, CXCL-10 and CXCL-11 in epithelial cancer cells [
203]. CD8+ T cells express CXCR3 and are recruited into the tumor by the CXCR3 ligands CXCL-9, CXCL-10, CXCL-11. The chemokines CXCL-9, 10 and 11 not only show angiostatic effects but are also important factors for the recruitment and activation of T helper 1 cells, which express the corresponding receptor CXCR3 [
203]. CTLs, therefore, have a great impact on the survival of CRC patients [
202]. Patients with MSI status show much higher CTL infiltration and a better prognosis than MSS patients [
204].CD4+ T helper cells (TH cells) differentiate into several subsets with divergent cytokine secretion and functions. TH cells modulate immune responses by activating or suppressing activities of immune cells such as macrophages, B cells and CTLs. CD4+ TH cells are divided by the ability to produce one or more signature cytokines and to express a lineage-specific transcription factor [
205].TH1 cells produce IFN-γ and TNF and express the Tbet transcription factor and the chemokine receptor CXCR3. Differentiation of naïve TH cells occurs in response to viruses and intracellular bacteria. After activation and differentiation, TH1 cells express the pro-inflammatory cytokines TNF and IFN-γ, which activate CTLs to kill infected cells, help B cells to present antigens and to activate CD8+ T cells more efficiently, and stimulate macrophages to phagocyte dead cells and debris. In CRC, the infiltration of TH1 cells is positively correlated with good clinical outcome [
44]. TH2 cells produce IL-4, IL-5 and IL-13 and express the transcription factor GATA-3 in response to extracellular pathogens. TH2 cells express the chemokine receptors CCR3, CCR4 and CCR10 that bind various CCL chemokines [
206]. TH2 lymphocytes polarize macrophages toward a M2 phenotype, thus, acting as tumor-promoting players. IL-4 promotes tumor proliferation and increases the production of ROS. In experimental mouse models of CRC, IL-4 and IL-4R-deficient mice develop fewer tumors than the control animals [
207]. This is a contradictory result as in CRC patients, the type 2 signature with IL-4, IL-5 and IL-13 has no prognostic advantage [
112].TH9 lymphocytes produce IL-9 and express PU.1. The role of TH9 cells in CRC is not clear as IL-9 has a strong inflammatory activity in experimental colitis, leading to colitis-associated cancer [
208]. The expression of IL-9, however, was significantly lower in patients with CRC than in the control tissue and correlated with staging and prognosis [
209]. TH17 lymphocytes produce IL-17A, IL-17F and IL-22 and express the transcription factor RORγt. TH17 lymphocytes express the chemokine receptors CCR6 and CXCR4 [
43]. As explained in the previous paragraph, TH17 cells are important for the protection against extracellular bacterial infection. In a mouse model of colorectal cancer, the inhibition of IL-17 leads to significantly reduced tumorigenesis [
109]. IL-17A induces the production of growth factors that stimulate the proliferation of myeloid cells, especially neutrophils. At the same time, IL-17A induces the production of CXCL-1 and CXCL-5, leading to the recruitment of the expanded myeloid cells into the tumor where they help to establish a tumor promoting microenvironment. Additionally, IL-17 promotes the production of IL-6 and TNF. IL-6, TNF and IL-17A promote the growth of CRC cells via STAT3 and NF-κB [
210]. The cytokine TGF-β induces the production of IL-22 in IL-17-positive TH17 cells and, subsequently, tumor progression [
211]. TH17 cells occur much more frequently in MMR proficient CRC tumors than in MMR-deficient tumors [
212].Regulatory T cells produce TGF-β, IL-10 and IL-35 and express the transcription factor forkhead box P3 (FOXP3) and the chemokine receptors CCR4, CCR5, CCR10 and CXCR3 [
205,
213]. The cytokine TGF-β induces in the TME the transcription factor FOXP3 and the differentiation of Tregs. In the TME, TAMs and MDSCs secrete CCL-17, CCL-22, CCL-5, CCL-6 or CCL-28 to recruit regulatory T cells. T regs show a high immune suppression capacity, employing more than a dozen suppression mechanisms. They can suppress immune cells by producing the cytokines IL-10, TGF-β or IL-35. In addition, they show suppressive activities by depleting soluble and membrane bound molecules needed for effector functions such as depletion of extracellular adenosine triphosphate (ATP) or the stripping of co-stimulatory molecules from the surface of DCs [
214,
215]. Although Tregs effectively suppress the activities of CTLs and NK cells, their role in sporadic cancer is still unclear. Some studies described a reduced overall survival in the presence of high frequencies of Tregs among tumor-infiltrating lymphocytes [
213,
216]. However, high levels of infiltrated Tregs were also associated with good prognosis [
217,
218].Natural killer (NK) cells belong to the family of innate lymphoid cells. NK cells are capable of killing tumor cells without antigen presentation and release TNF and IFN-γ. In CRC, the infiltration of cytotoxic NK cells is associated with a better prognosis [
219].
Chronic inflammation is characterized by mucosal infiltration of inflammatory immune cells that interact with the local microbiota and induce a pro-inflammatory milieu.
Regulatory B cells (Bregs) are immune suppressive and reduce the tissue damaging TH1/TH17 response by secreting IL-10 and IgA antibodies and restore the cytokine-induced balance [
220]. Several cytokines, including IL-6, IL-33 and IL-35, induce the transition of B cells to Bregs [
221]. Breg cells suppress the anti-tumorigenic immune response and lead to tumor progression by the tumor-promoting cytokine IL-10 [
222]. In CRC, this tissue preserving and TH1-reducing activity mediated by IL-10 is one of the escape mechanisms of colorectal cancer cells leading to excessive proliferation of the tumor.Other central players in coordinating immune responses against invading pathogens are dendritic cells. DCs are a heterogeneous population of antigen presenting cells that scan the environment for danger signals and antigens. In homeostasis, immature DCs have the capacity to process and present antigens to primed T cells. To avoid tissue damage and exacerbating immune responses these immature DCs secrete, similar to regulatory B cells, the immune-suppressive cytokines IL-10 and TGF-β and show low expression of co-stimulatory molecules. In the presence of PAMPs or DAMPs and inflammatory cytokines, the maturation process starts. Mature DCs upregulate co-stimulatory molecules and secrete the pro-inflammatory cytokines IL-12, IL-6, TNF, IL-1β that are required for priming, activation and proliferation of T cells. Several tumor-derived cytokines such as TGF-β, TNF, IL-6 or IL-10 are able to prevent the maturation of DCs, the infiltration of mature DCs, the presentation of antigens and the activation of T and NK cells [
223,
224].The CMS classification made clear that CRC is a very heterogenous disease, especially regarding the tumor microenvironment. As an effective anti-tumorigenic response depends on infiltrating effector immune cells of the innate and adaptive immune system, chemokines that facilitate the recruitment of these cells are of great importance. In the CMS2 and the CMS3 subtypes, there is hardly any immune cell infiltration; therefore, these subtypes show poor anti-tumor immunity. One good example for the pivotal role of some chemokines is the expression of CXCR3 chemokines in myeloid cells. The pro-inflammatory cytokine IFN-γ induces CXCR3 chemokines, e.g., CXCL-9 and CXCL-10, as part of the interferon-stimulated gene cluster that then attracts T cells into the tumor [
225]. However, as shown by CMS4, the infiltration of immune cells is not enough to fight the tumor. Cytokines within the TME determine the balance between tumor-promoting and tumor-destructive features of the immune cell infiltrate and the cells of the microenvironment (
Figure 4). Cytokine and chemokine networks that are shaped by inflammatory processes regulate this balance. In the context of CRC, this is further complicated by the genotype of the tumor. Both MMR-deficient and -proficient CRCs show strong stromal remodeling with reduced amounts of fibroblasts compared with healthy colon samples. Both CRC types are characterized by inflammatory interaction networks of malignant epithelial cells, monocytes, CAFs and TANs at the luminal margin of the tumors. In both CRC subtypes, the pro-inflammatory cytokines IL1-β, IL-6 and TNF upregulate the chemokines CXCL-1/2/3/5/6/8 in CAFs, monocytes and cancer cells; this attracts CXCR1/2-positive neutrophils to the tumor. MMR-deficient CRCs respond to immune checkpoint therapy, whereas MMR-proficient CRCs are unresponsive to immunotherapy. The difference between both tumor types are foci in the MMR-deficient tumors with IFN-γ-positive and CXCL-13-positive T cells that form positive feedback loops to attract more activated T cells [
226].


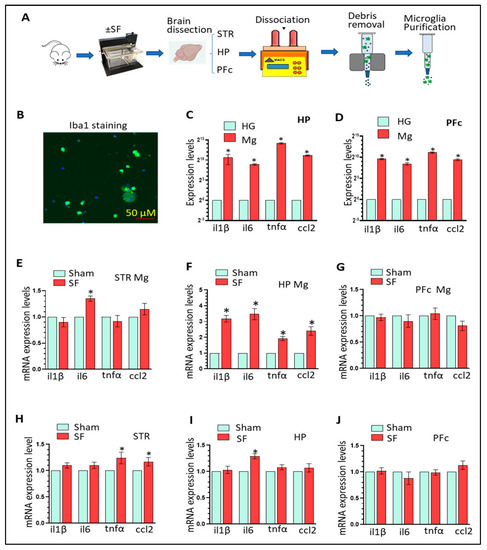
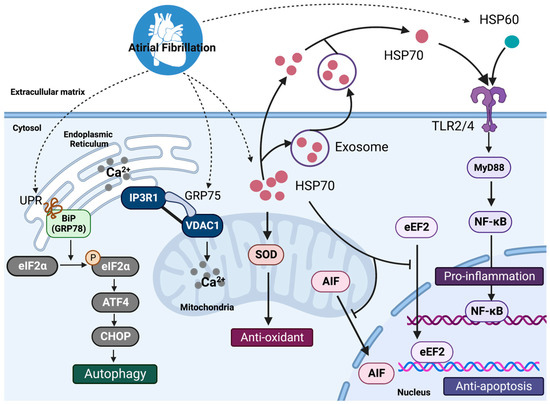

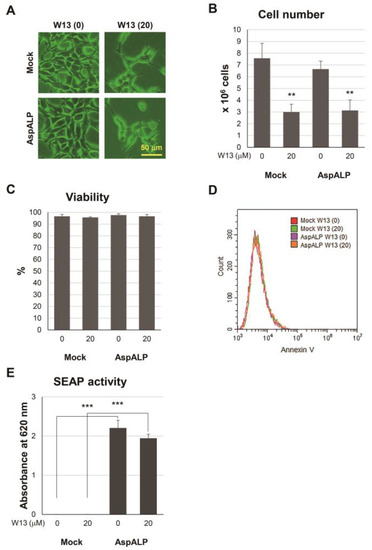
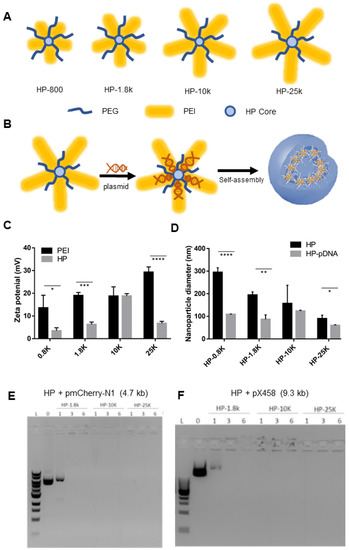
留言 (0)