The extracellular matrix (ECM) of a multicellular eukaryotic organism not only provides a stable scaffold that keeps cells in appropriate place and orientation, but is also involved in signaling mechanisms that regulate growth and differentiation patterns, cell homeostasis, and pathogen defense [
1,
2,
3,
4]. For the study of the ECM and other essential components required for multicellularity, the model organism Volvox carteri (Chlamydomonadales) and its close relatives, the volvocine algae, lend themselves as target organisms. These volvocine algae are particularly suitable for investigating the evolutionary transition to multicellular life because they diverged relatively recently from unicellular relatives, and extant species display a range of size, cell numbers and organizational complexity, from unicellular and colonial genera to multicellular genera with full germ-soma division as found in Volvox [
5,
6,
7,
8,
9,
10,
11,
12,
13]. In the course of evolution to multicellularity, the development of a complex, multifunctional ECM from the simple cell wall of a unicellular ancestor was required. In volvocine algae, a correlation between the complexity of the organisms and the proportion of ECM per organism can be observed: A 200 nm thick ECM layer surrounds the plasma membrane of the unicellular alga Chlamydomonas reinhardtii (mostly called ‘cell wall‘), which accounts for about 10% of the total volume of the cell. In the multicellular alga Volvox carteri, ECM surrounds not only the individual cells, but the cells form a spherical monolayer that encloses ECM structures and, in addition, the cell monolayer is surrounded externally by ECM layers (
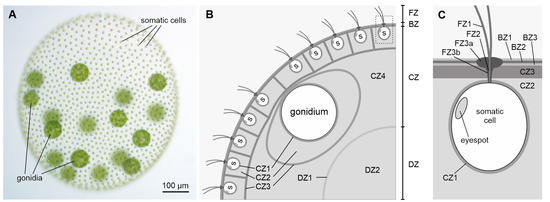
Figure 1).In an adult Volvox alga, the volume fraction of the ECM thus accounts for more than 95% of the organism. Despite the general simplicity of Volvox with only two cell types, its ECM is surprisingly elaborated, consisting of many region-specific, anatomically distinct structures, which are modified under physiological, metabolic, or developmental control [
8,
14]. There are four main zones of the Volvox ECM, the flagellar zone (FZ), the boundary zone (BZ), the cellular zone (CZ) and the deep zone (DZ), which are further subdivided into subzones (
Figure 1) [
14]. The FZ comprises specializations of the ECM around the flagella, with the so-called flagellar hillock (FZ3a) and the flagellar tunnel wall (FZ3b) together forming an ECM tube at the basal end of each flagellum. The BZ contains portions of the ECM that, except in periflagellar regions, are continuous over the surface of the organism and are not structurally continuous with deeper layers. The CZ1 immediately surrounds the plasma membrane of all somatic and reproductive cells. The particularly robust CZ1 around each reproductive cell (gonidium) is also called ‘gonidial vesicle’. When the gonidia begin to divide, the cell membranes detach from the gonidial vesicles, but the gonidial vesicles remain intact, and all further development to juveniles occurs inside the gonidial vesicles. CZ3 is a coherent, more fibrous material that forms honeycomb-like chambers at greater distances around individual cells. CZ2 consists of rather amorphous ECM components that fill the space between CZ1 and CZ3. CZ4 characterizes the unstructured, very loose material that lies within the somatic cell layer but outside the DZ and in which the gonidia are embedded. The DZ comprises ECM components below the CZ. DZ2 is poorly structured, fills the deepest region of the spheroid and is by far the largest region. The structure of DZ2 is similar to that of CZ4. DZ1 is a thin, fibrous boundary layer that surrounds DZ2. At the time when the matured juveniles hatch from their mother spheroid, they not only have to come out of the gonidial vesicle, but birth canals must also be formed through the mother spheroid [
5]. To achieve this, not only the gonidial vesicle but also the ECM of the mother spheroid directly above the juveniles must be removed, while the ECM of the hatching juveniles must remain intact and completely unharmed. At least two lytic enzymes appear to be important for ECM degradation during the hatching process in V. carteri: VheA [
15] and Lsg2 [
16]. In addition, the presence of a certain pherophorin, PhS [
17,
18,
19], in the maternal CZ1 seems to be required as an ECM plasticizer [
19]. In a synergistic action, the ECM is first softened and then dissolved so that the juveniles can hatch. It has long been known that the ECMs of volvocine algae are mainly composed of hydroxyproline-rich glycoproteins (HRGPs) [
20]. These HRGPs not only dominate the ECM composition of green algae, but also represent a main constituent of the ECMs of embryophytic land plants [
8,
18,
21,
22,
23]. In both V. carteri and C. reinhardtii, a number of ECM components have been characterized in more detail [
8,
19,
24,
25,
26,
27,
28,
29,
30,
31]. Many of the ECM components characterized in V. carteri are HRGPs, which appear to be responsible for the assembly, architecture, and structure of the multilayered ECM [
8,
17,
28,
29,
32,
33,
34,
35,
36,
37,
38,
39]. The ECM is subject to change in the course of development. This is particularly evident during the switch from vegetative to sexual development, which is triggered by the V. carteri sex-inducer and leads to the synthesis of alternative ECM components [
8,
28,
29,
32,
34,
35,
39,
40,
41]. Because the ECM expands dynamically and also needs to be remodeled during development, proteases and other lytic enzymes also exist in the ECM [
8,
27,
42]. Lytic enzymes are also required for local degradation of the maternal ECM during hatching of the daughter spheroids [
15,
16,
19]. In addition, there are enzymes in the ECM that serve to mobilize sulfates and phosphates [
43,
44]. Finally, defense enzymes have been found in the ECM, e.g., chitinases against fungi [
45]. Furthermore, using the sequences of known ECM glycoproteins, analyses of the sequenced genomes of V. carteri [
46] and C. reinhardtii [
47] identified additional putative ECM proteins [
19]. One long-known protein family with importance for ECM biosynthesis stands out among all the previous studies not only because of the number of family members: the pherophorins [
17,
28,
33,
34,
35,
36]. In V. carteri, this protein family comprises 118 members [
19] including the V. carteri sex-inducer, which constitutes a pherophorin-related protein [
34]. In C. reinhardtii 35 members of the pherophorin family could be identified [
48]. Pherophorins typically have a dumbbell-like domain structure with two globular domains separated by a rod-shaped, highly proline-rich domain with a strongly varying length [
8,
17,
18,
35]. The prolines of the rod-shaped domain are post-translationally modified to hydroxyproline [
8,
28]. All pherophorins biochemically studied so far were glycoproteins, i.e., they contained covalently-linked oligosaccharide chains (glycans). The building blocks of these glycans are predominantly the sugars arabinose, galactose and mannose, indicating O-linked glycosylation of serine, threonine and hydroxyproline [
8,
18,
28]. However, in the rod-shaped domain, O-glycosylation is primarily enabled by the abundant hydroxyprolines. The O-linked sugars of the pherophorins are highly sulfated [
32,
49] and unusual phosphodiester linkages between two arabinose residues have also been identified [
17,
36,
50]. These phosphodiester bridges could be responsible for intermolecular cross-links between the polysaccharide parts of pherophorins [
36]. For two purified pherophorins, pherophorin-DZ1 and pherophorin-DZ2, polymerization into an insoluble fibrous network was demonstrated in vitro [
36], although the chemistry behind the polymerization remained unclear.Among the pherophorins studied so far, distinctly different expression patterns were found. While some pherophorins showed constitutive expression from the end of embryogenesis [
34,
51], expression of others was strongly enhanced after induction of sexual reproduction [
17,
28,
34,
35,
36] or, in others, detectable only at specific developmental stages [
37]. In a whole transcriptome RNA-Seq analysis, 84% of the V. carteri pherophorins examined showed cell type-specific expression, and the number of cell type-specific pherophorins was similar for each of the two cell types [
52]. However, all pherophorins of V. carteri that were extracted from the ECM and characterized, were specifically expressed by somatic cells. Thus, all previous biochemical characterizations of individual pherophorin proteins referred only to pherophorins synthesized by the somatic cell type; not a single pherophorin studied was expressed by gonidia.The first pherophorin isolated from the ECM was SSG185 [
33], although it was not clear at the time that it was a member of a large protein family. In particular, the protein chemical nature and posttranslational modifications of SSG185 were studied in detail [
8,
18,
28,
32,
33,
49,
50,
53]. In the process, it was also shown that SSG185, as with most of the pherophorins studied later, possesses a hydroxyproline-rich domain (HR domain) [
33]. For in-situ immunolocalization of SSG185, the researchers used a part of SSG185 containing the HR domain for antibody production. The latter is also present in other pherophorins. Therefore, the previous, antibody-based finding that SSG185 localizes to CZ3 and forms the honeycomb-like cellular compartments of the V. carteri ECM [
33], needs review, especially since several later publications built on this finding [
5,
8,
17,
18,
35,
50].
In this study, we present a phylogenetic analysis of all pherophorin-related genes found in V. carteri and C. reinhardtii. We are investigating the HR domains of pherophorins and, for pherophorins from V. carteri, we also disclose whether they are expressed in a cell type-specific manner and, if so, in which cell type. We then show the precise localization of two pherophorins by live-cell imaging using generated transformants that stably express fluorescence-tagged pherophorins. For this, we have selected the aforementioned pherophorin SSG185, a pherophorin that is mainly expressed by somatic cells. In addition, we localized a pherophorin that is mainly expressed by gonidia, namely pherophorin PhG. The results provide insights into the evolution of the pherophorin protein family, show distinctly different properties of soma-specific and gonidia-specific pherophorins, and provide information on how somatic cells and gonidia cooperatively assemble the ECM of V. carteri.



留言 (0)