
記住我
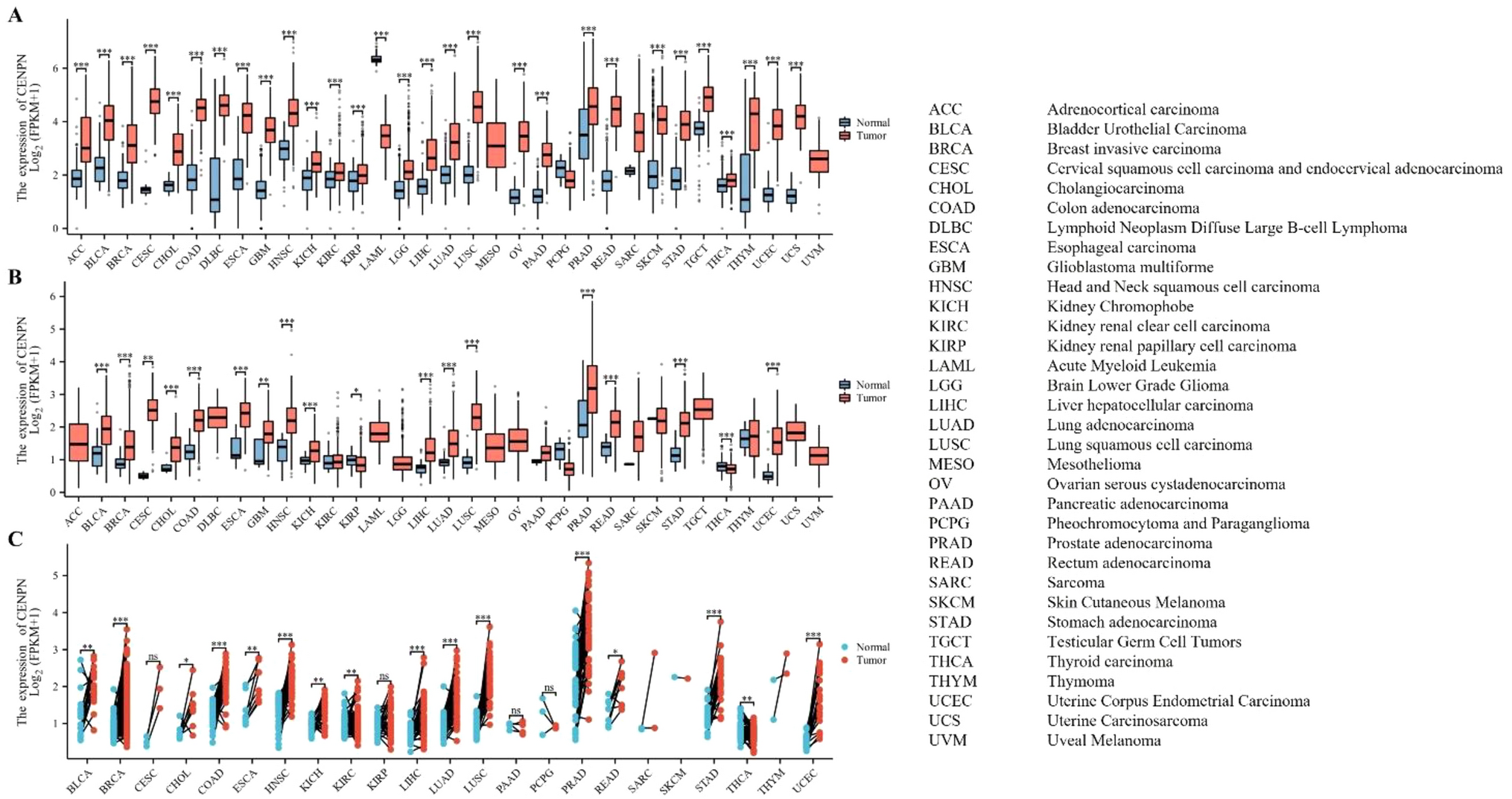



CENPN expression was analyzed in 33 cancer types (Table 1). The unpaired sample analysis, which included 18,102 samples, revealed low CENPN expression in LAML (P < 0.001) and high expression in ACC, BLCA, BRCA, CESC, CHOL, COAD, DLBC, ESCA, GBM, HNSC, KICH, KIRC, KIRP, LGG, LIHC, LUAD, LUSC, OV, PAAD, PRAD, READ, SKCM, STAD, TGCT, THCA, THYM, UCEC, and UCS (all P < 0.001) compared to normal samples (Fig. 1A). MESO and UVM were excluded due to insufficient normal samples. The unpaired analysis, involving 11,123 samples, showed significantly higher CENPN expression in BLCA, BRCA, CHOL, COAD, ESCA, HNSC, KICH, LIHC, LUAD, LUSC, PRAD, READ, STAD, UCEC (all P < 0.001), CESC (P = 0.003), and GBM (P = 0.009) relative to paracancerous tissue (Fig. 1B). ACC, DLBC, LAML, LGG, MESO, OV, TGCT, UCS, and UVM were not analyzed due to insufficient paracancerous samples, while no significant differences were observed in KIRC, PAAD, PCPG, SARC, SKCM, and THYM (P > 0.050). The paired sample analyses, involving 1404 samples from 23 cancers and their corresponding paracancerous samples, demonstrated significant increases in CENPN expression in BRCA, COAD, HNSC, LIHC, LUAD, LUSC, PRAD, STAD, UCEC (all P < 0.001), BLCA (P = 0.002), CHOL (P = 0.023), ESCA (P = 0.008), KICH (P = 0.004), KIRC (P = 0.008), and READ (P = 0.012) (Fig. 1C). These findings highlight the differential expression of CENPN across various cancer types, setting the stage for a deeper exploration of its diagnostic and prognostic value.
Table 1 TCGA cancer abbreviations and the corresponding cancer typeFig. 1
The expression of CENPN mRNA in pan-cancer. A Expression of CENPN between the 33 cancers and normal tissues in unpaired sample analysis; B Expression of CENPN between the 33 cancers and paracancerous tissues in unpaired sample analysis; C Paired sample analysis of CENPN mRNA expression between 23 cancers and paracancerous tissues in BLCA, BRCA, CESC, CHOL, COAD, ESCA, HNSC, KICH, KIRC, KIRP, LIHC, LUAD, LUSC, PAAD, PCPG, PRAD, READ, SARC, SKCM, STAD, THCA and UCEC. ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001. ns, Not Significant
3.2 Diagnostic value of CENPN across cancersThe diagnostic value of CENPN was significant across various cancers, as shown in Fig. 2A–P. In 23 cancers, the AUC for CENPN exceeded 0.6, and in 16 cancers, it surpassed 0.8, indicating high diagnostic value. These cancers included ACC (AUC = 0.848), BRCA (AUC = 0.817), COAD (AUC = 0.954), CHOL (AUC = 0.863), DLBC (AUC = 0.847), ESCA (AUC = 0.917), GBM (AUC = 0.843), HNSC (AUC = 0.922), LUAD (AUC = 0.845), LUSC (AUC = 0.988), LIHC (AUC = 0.844), OSCC (AUC = 0.915), PCPG (AUC = 0.810), STAD (AUC = 0.923), SARC (AUC = 0.894), and UCEC (AUC = 0.941) (Supplementary Fig. 1). The strong diagnostic performance of CENPN prompted further investigation into its prognostic implications across cancers.
Fig. 2
Receiver Operator Characteristic (ROC) curve of CENPN in 16 Cancers. Cancers with AUC > 0.8 for CENPN: (A) ACC, (B) BRCA, (C) COAD, (D) CHOL, (E) DLBC, (F) ESCA, (G) GBM, (H) HNSC, (I) LUSC, (J) LUAD, (K) LIHC, (L) OSCC, (M) PCPG, (N) STAD, (O) SARC, (P) UCEC
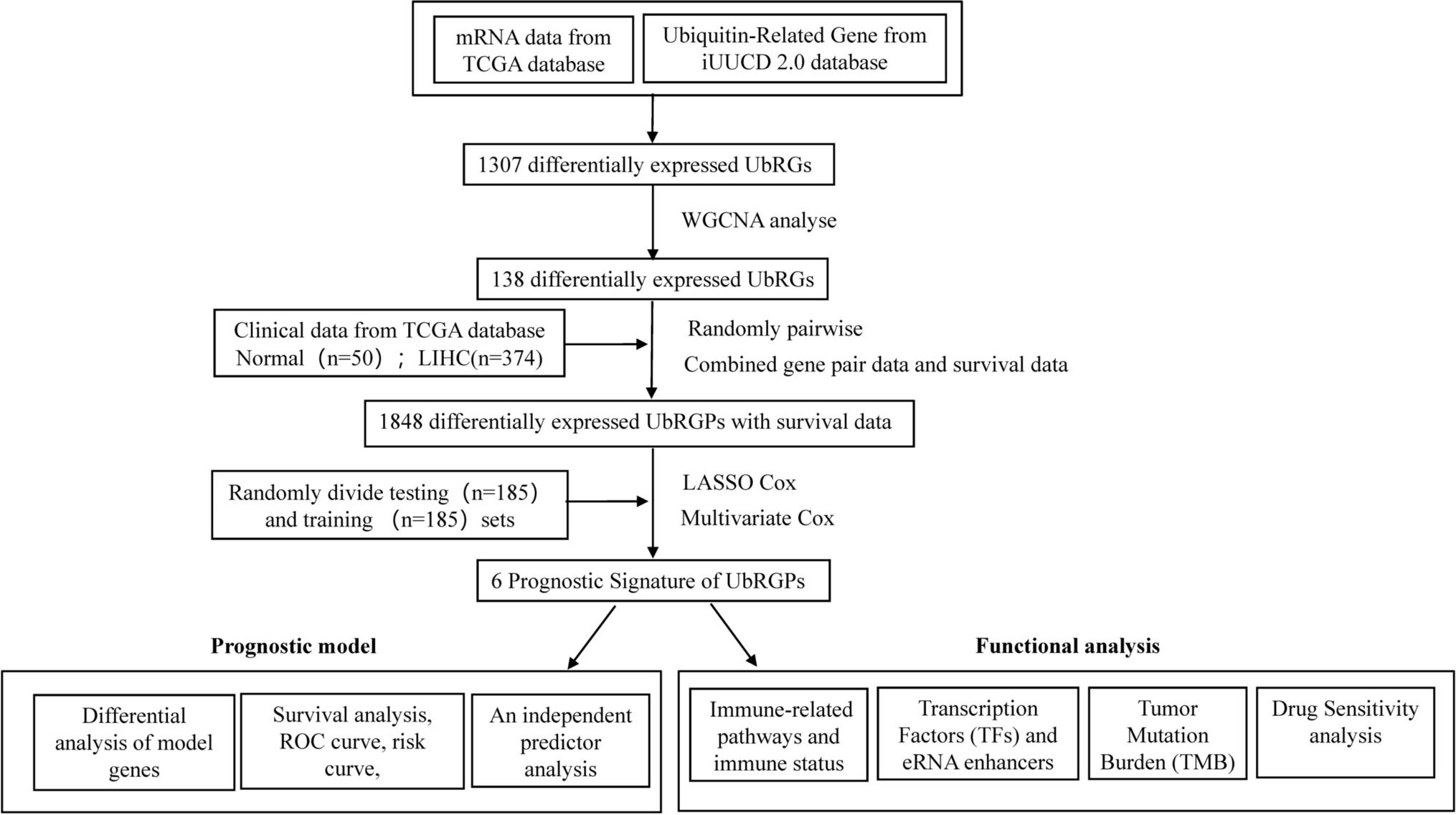
3.3 Prognostic value of CENPN across cancersOur results demonstrate that CENPN expression levels are significantly associated with patient outcomes across several cancer types (Fig. 3). Low CENPN expression was significantly linked to better OS in ACC, BRCA, HNSC, KICH, KIRP, LAML, LIHC, LUAD, LGG, MESO, OSCC, PAAD, SARC, and THCA, while high CENPN expression was significantly associated with better OS in THYM and OV (Fig. 4A–P). Regarding DSS, higher CENPN expression levels were associated with better survival outcomes in OV but correlated with poorer outcomes in ACC, BLCA, BRCA, ESAD, KICH, KIRC, KIRP, LGG, LIHC, LUAD, MESO, and PAAD (Fig. 3 and Supplementary Fig. 2A-M). Similarly, in the PFI analysis, higher CENPN expression was correlated with shorter PFI in ACC, BLCA, BRCA, KIRC, LGG, LIHC, LUAD, MESO, PAAD, PCPG, and THCA (Fig. 3 and Supplementary Fig. 3A-O). These associations suggest that CENPN expression may serve as a potential marker for predicting disease progression and recurrence.
Fig. 3
K-M analysis for high- and low- CENPN gene expression in cancers. Forest plot of CENPN OS in 16 cancers (A), DSS in 13 cancers (B), and PFI in 15 cancers (C)
Fig. 4
Correlations between CENPN and prognosis in 16 cancers. OS K-M curve for CENPN in 16 cancers. The unit of X-axis is month. (A) ACC, (B) BRCA, (C) HNSC, (D) KICH, (E) KIRP, (F) LAML, (G) LIHC, (H) LUAD, (I) LGG, (J) MESO, (K) OSCC, (L) OV, (M) PAAD, (N) SARC, (O) THCA, (P) THYM
3.4 CENPN expression in different immune and molecular subtypes across cancersOur analysis revealed that variations in CENPN expression significantly affect overall survival (OS) in 16 cancer types. Further investigation of these and an additional 17 cancer types showed significant differences in CENPN expression across both immune and molecular subtypes. In nine of the 16 cancers, significant variations in CENPN expression among immune subtypes were observed, including ACC (six subtypes), BRCA (five subtypes), KIRP (six subtypes), LGG (four subtypes), LIHC (five subtypes), LUAD (five subtypes), OV (four subtypes), PAAD (five subtypes), and SARC (five subtypes) (Fig. 5A–I). Moreover, notable differences in CENPN expression across molecular subtypes were found in six cancer types: ACC, BRCA, KIRP, LGG, LIHC, and OV (Fig. 6A–F). For the remaining 17 cancer types, significant variance in CENPN expression was observed in immune subtypes (BLCA, COAD, ESCA, KIRC, LUSC, PRAD, READ, STAD, TGCT, UCEC, and UVM) (Supplementary Fig. 4A–K) and molecular subtypes (COAD, ESCA, LUSC, PCPG, PRAD, SKCM, STAD, and UCEC) (Supplementary Fig. 5A–H). Having established the subtype-specific variations in CENPN expression, we next examined its correlation with clinical phenotypes.
Fig. 5
Correlations between CENPN expression and immune subtypes in 9 cancers. (A) ACC, (B) BRCA, (C) KIRP, (D) LGG, (E) LIHC, (F) LUAD, (G) OV, (H) PAAD, (I) SARC. C1 (wound healing), C2 (IFN-γ dominant), C3 (inflammatory), C4 (lymphocyte deplete), C5 (immunologically quiet), and C6 (TGF-β dominant)
Fig. 6
Correlations between CENPN expression and molecular subtypes in 6 cancers. (A) ACC, (B) BRCA, (C) KIRP, (D) LGG, (E) LIHC, (F) OV
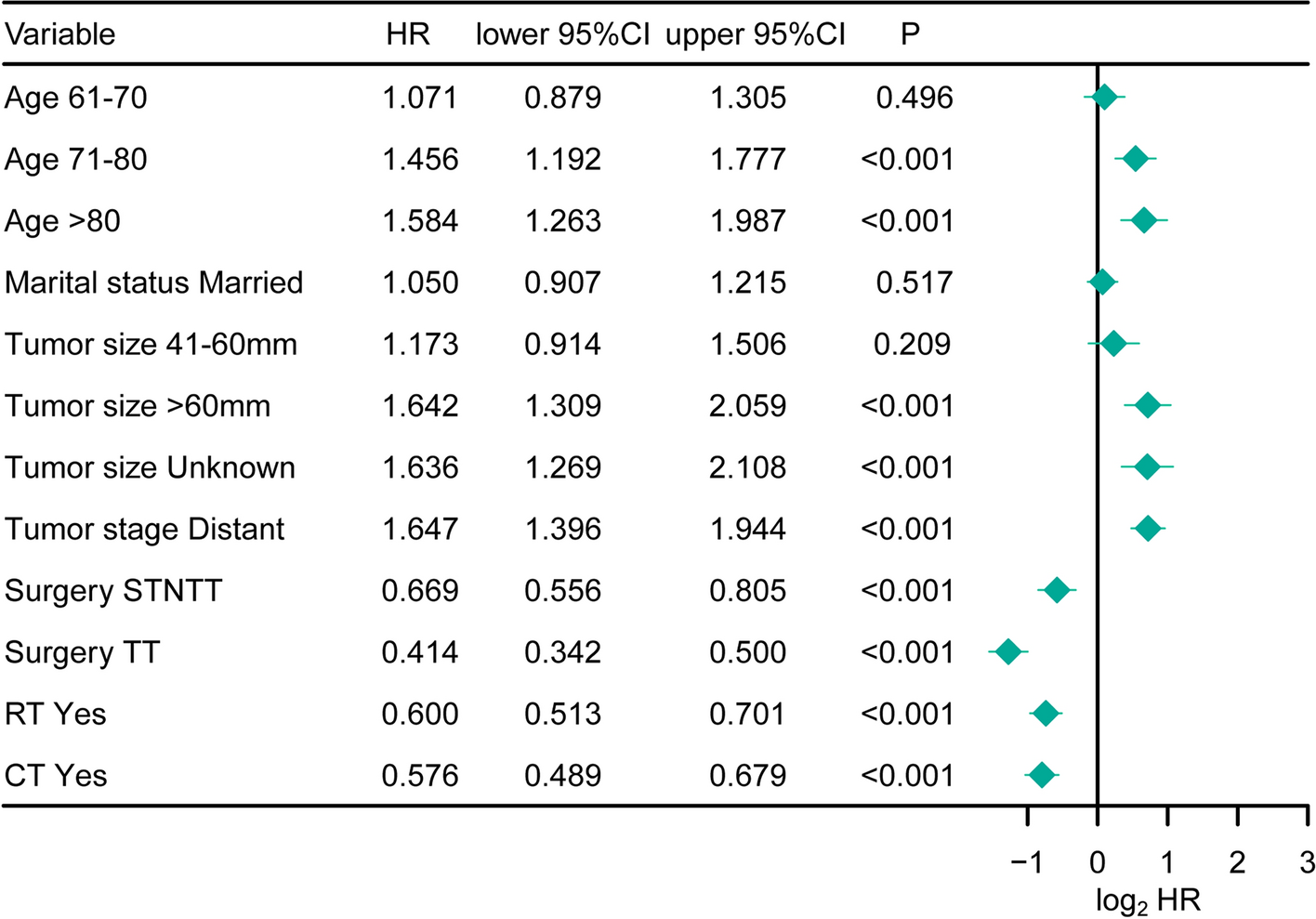
3.5 Correlation of CENPN expression with clinical phenotypes in various cancersThe analysis of CENPN expression in relation to age across various tumor types revealed higher expression levels in patients aged ≤ 60 years with BRCA (Fig. 7A; P < 0.001), ESAD (Fig. 7B; P = 0.002), ESCC (Fig. 7C; P = 0.003), and THYM (Fig. 7G; P < 0.001). Additionally, patients aged ≤ 65 years with LUSC (Fig. 7E; P < 0.001) exhibited higher expression levels, while those with STAD > 65 years showed higher CENPN expression (Fig. 7F; P = 0.005). In LGG patients aged > 40 years (Fig. 7D; P = 0.002), higher expression levels were observed. However, no significant age-related correlations were found in other cancer types. Investigation of tumor stage relevance revealed significantly increased CENPN expression in the early tumor stage of 15 cancer types (Fig. 8), including BLCA, BRCA, CESC, CHOL, COAD, ESCA, HNSC, KICH, KIRC, LIHC, LUAD, LUSC, OSCC, STAD, and THCA, suggesting the potential clinical value of CENPN in the early diagnosis of these tumors. Given the clinical relevance of CENPN expression, we proceeded to explore its genetic alterations in various cancers.
Fig. 7
Association between CENPN expression and age in (A) BRCA, (B) ESAD, (C) ESCC, (D) LGG, (E) LUSC, (F) STAD and (G) THYM
Fig. 8
Association between CENPN expression and tumor stage. *P < 0.05, **P < 0.01, ***P < 0.001. ns, not statistically significant
3.6 Genetic alteration of CENPNThe genetic mutations in CENPN expression across various cancers were evaluated using the cBioPortal online tool. The analysis included 32 studies, comprising 10,967 samples from the TCGA PanCancer Atlas. Within the amino acid range of 0 to 339, a total of 66 mutation sites were identified, including 52 missense mutations, 12 truncating mutations, one splicing mutation, and one SV/fusion mutation. The S107L mutation site was the most frequently observed (Fig. 9A). The primary mutation types were deep deletion, missense mutation, and amplification, with CENPN mutations being most prevalent in PRAD, UCEC, BLCA, COAD, BRCA, OV, SKCM, and ESCA (Fig. 9B). Across the 32 cancer types, shallow deletion was a common occurrence in CENPN expression, except in ACC, KICH, and PCPG (Fig. 9C). These genetic insights prompted us to further explore the functional implications of CENPN through protein interaction and enrichment analyses.
Fig. 9
Genetic alteration of CENPN across cancers. A Mutation diagram of CENPN across protein domains; B Bar chart of CENPN mutations across cancers studies based on TCGA PanCancer Atlas Studies; C Mutation counts and types of CENPN across cancers
3.7 The PPI, functional enrichment, and gene set enrichment of CENPN in cancersFifty closely associated genes with CENPN were retrieved from STRING, and a PPI network was constructed using the specified threshold (Fig. 10A). The top 14 hub genes were identified: H4C15, H4C12, H4-16, H4C8, H4C1, H4C2, H4C11, H4C14, H4C9, H4C4, CENPN, H4C13, H4C3, and H4C5 (Fig. 10B). These hub genes exhibit significant associations in cancers where CENPN expression influences prognosis, except in KICH, LAML, MESO, and OV (Fig. 10C). GO/KEGG enrichment analyses were performed to investigate the functional roles of these genes. The primary GO terms for biological processes (BP) included protein-DNA complex subunit organization, protein-DNA complex assembly, and nucleosome assembly. Cellular components (CC) encompassed chromosomes, centromeric regions, and nucleosomes, while molecular functions (MF) involved protein heterodimerization activity. The top KEGG pathways included neutrophil extracellular trap formation, alcoholism, and systemic lupus erythematosus (Fig. 10D). Notably, in cancer scenarios, CENPN activates pathways associated with the cell cycle, apoptosis, EMT, DDR, the TSC/mTOR pathway, and the hormone AR, while inhibiting pathways related to RAS/MAPK signaling, the hormone ER, RTK, PI3K/AKT, and the hormone AR (Fig. 10E).
Fig. 10
The PPI network and functional enrichment analysis of CENPN. A The PPI network of CENPN, B The top 14 hub genes of PPI network, C The association hub gene with CENPN in 16 cancers present as heatmap. ∗P < 0.05, ∗∗P < 0.01, D GO/KEGG pathway enrichment for CENPN and closed interact genes, E CENPN with pathway activity or inhibition
GSEA results across 16 cancer types (Fig. 11A–P) revealed frequently enriched pathways, including the regulation of DNA replication, chromosome separation, sister chromatid segregation, nuclear chromosome segregation, antigen binding, and immunoglobulin complex circulation. These observations suggest a strong association between CENPN and processes such as DNA strand replication, cell cycle regulation, recombination, and transcription.
Fig. 11
GSEA functional enrichment analysis of CENPN expression in 16 cancers. The top 5 GSEA functional enrichment pathways of CENPN in (A) ACC, (B) BRCA, (C) HNSC, (D) KICH, (E) KIRP, (F) LAML, (G) LGG, (H) LIHC, (I) LUAD, (J) MESO, (K) OSCC, (L) OV, (M) PAAD, (N) SARC, (O) THCA, (P) THYM. The Y-axis represents one gene set and the X-axis is the distribution of logFC corresponding to the core molecules in each gene set
3.8 Single-cell function evaluation of CENPNThe functional role of CENPN in various cancer types was investigated using CancerSEA, which facilitated the analysis of CENPN's association with multiple functional states of cancer cells at the single-cell level. The findings revealed a positive correlation between CENPN expression and activities related to cell cycle regulation, DNA repair, DNA damage response, cell invasion, and cell proliferation. Conversely, negative correlations were observed between CENPN expression and conditions of hypoxia, cellular quiescence, and angiogenesis, although these associations were relatively weak (Fig. 12A). Furthermore, the relationship between CENPN and functional statuses in specific cancer types was examined. The results showed that CENPN was positively correlated with DNA damage in Acute Lymphoblastic Leukemia (ALL) and with cell cycle regulation and cell invasion in Acute Myeloid Leukemia (AML). In Breast Cancer (BRCA), associations with cell cycle regulation, cell proliferation, DNA repair, and DNA damage were identified. However, in certain scenarios, CENPN exhibited negative correlations, particularly in Colorectal Cancer (CRC) with metastasis, hypoxia, and angiogenesis, and in Uveal Melanoma (UM) with DNA repair, apoptosis, invasion, and DNA damage. In Head and Neck Squamous Cell Carcinoma (HNSC), a negative correlation with quiescence was identified (Fig. 12B-L).
Fig. 12
The CancerSEA database’s single-cell functional analysis reveals the role of CENPN. (A) The interactive bubble chart present correlation of CENPN with functional state in 18 cancers. The correlation of CENPN with functional state in (B) acute lymphoblastic leukemia (ALL), (C) Acute myeloid leukemia (AML), (D) BRCA, (E) Colon and Rectal Cancer (CRC), (F) Retinoblastoma (RB), (G) GBM, (H) HNSC, (I) LUAD, (J) Prostate cancer (PC), (K) Renal cell carcinoma (RCC), (L) Uveal Melanoma (UM) X-axis represents different gene sets; ***P < 0.001, **P < 0.01, *P < 0.05.
3.9 Relationship between CENPN expression level and tumor immune cell infiltrationTo evaluate the association between CENPN, immune infiltration, and immune regulation, a heatmap featuring CENPN alongside markers for immune cells and factors was generated. The findings revealed that CENPN displayed a positive correlation with T helper, antigen-presenting dendritic cells (aDC), central memory T cells (Tcm), and Th2 cell infiltration across 33 cancers, but exhibited a negative correlation with the majority of other immune cell types (Fig. 13). Regarding immunostimulators, CENPN showed positive associations in BLCA, BRCA, KIRC, and THCA (Fig. 14A). Intriguingly, CENPN exhibited a negative correlation with various immunoinhibitors, particularly in COAD, ESCA, HNSC, PRAD, READ, and SKCM (Fig. 14B). For MHC molecules, a positive correlation between CENPN and most MHC molecules was observed in BLCA, BRCA, KIRC, LGG, and THCA, while a negative correlation was evident in ACC, GBM, HNSC, KICH, LUAD, LUSC, PRAD, SKCM, TGCT, and UCEC (Fig. 14C). Cytokines in BLCA, BRCA, KIRC, and THCA demonstrated predominantly positive correlations with CENPN, whereas negative correlations were observed in GBM, PRAD, and TGCT (Fig. 14D). Regarding cytokine receptors, CENPN was negatively correlated with the majority of them in ESCA, GBM, HNSC, LUSC, PRAD, READ, SKCM, and STAD, but displayed a positive correlation in KIRC and THCA (Fig. 14E).
Fig. 13
Relationship between CENPN expression and immune cell infiltration in different cancers. *P < 0.05, **P < 0.01, ∗∗∗P < 0.001
Fig. 14
Co-expression of CENPN and immune-related genes. (A) Immunostimulators, (B) Immunoinhibitors, (C) MHC molecules, (D) Chemokines, (E) Chemokine receptors. ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001
3.10 Correlation of CENPN expression with DNA Methylation.The UALCAN online tool was employed to investigate the promoter methylation levels of CENPN among different patient groups and normal individuals across various cancer types. The beta value, which ranges from 0 (unmethylated) to 1 (fully methylated), serves as an indicator of DNA methylation level. Different beta value cutoffs were considered to identify hypermethylation (0.7–0.5) or hypomethylation (0.3–0.25). As shown in Fig. 15, the methylation level of the CENPN promoter is significantly lower in most tumor groups compared to the normal group (all P < 0.010).
Fig. 15
The promoter methylation level of CENPN in cancers
Based on data from the CTD database, CENPN is associated with 66 chemicals, 42 of which can upregulate CENPN and 24 that can downregulate it (Table 2). Additionally, the top 20 relationships between CENPN and other genes via chemical associations were identified. The results showed that CENPN has a high correlation with inner centromere protein (INCENP), non-SMC condensin II complex subunit G2 (NCAPG2), timeless circadian regulator (TIMELESS), structural maintenance of chromosomes 2 (SMC2), and centromere protein H (CENPH) (Table 3).
Table 2 Interacting chemicals of CENPN from CTDTable 3 Relationship of CENPN with genes via chemical interaction, based on the CTD database
留言 (0)