
記住我
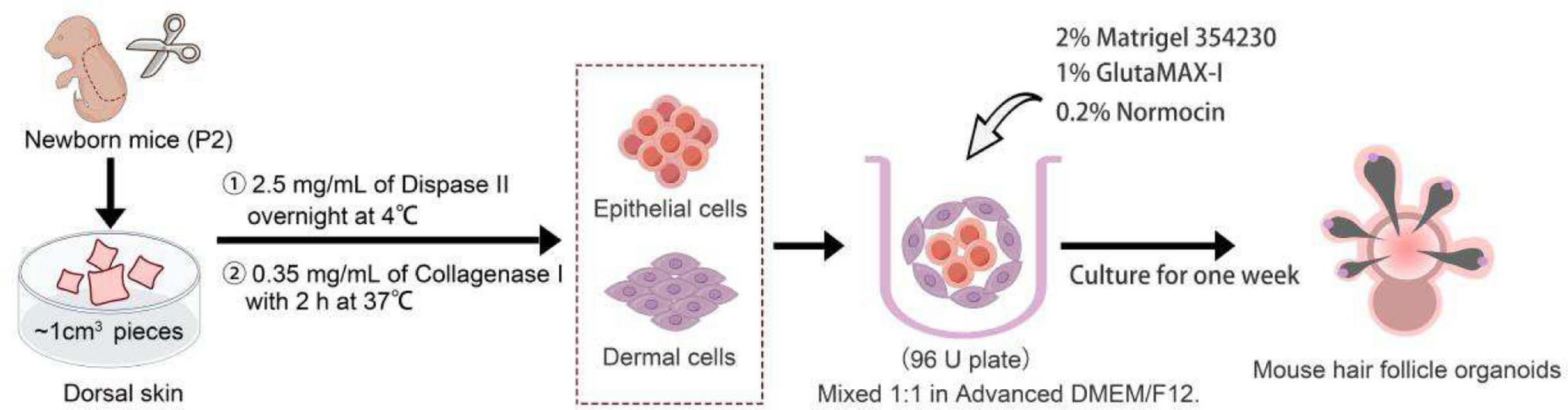
The formation of HF germs during development initiates HF induction through epithelial-mesenchymal interactions. In this study, we isolated the dermal and epidermal layers from the skin of neonatal mice and conducted HF reconstitution in Advanced DMEM/F12 supplemented with 2% Matrigel (Fig. 1 and S1). As shown in Fig. 2A, on the day 4, the dermal-epidermal aggregates had formed cyst-like structures composed of epidermal precursors enveloped by dermal cells; these structures are essential for self-organization and for mimicking the cell-to-cell signaling processes involved in HF development. On the day 9, vigorous and healthy HFs were observed. However, by the second to third week, the HFs lost vitality, gradually underwent atrophy, and completed their entire life cycle. On the fourth day, prior to the formation of HF buds, the mHFOs were transferred to OMM for further culture (Fig. 2B and C). OMM consists of Advanced DMEM/F12 and Neurobasal media in a 1:1 ratio, supplemented with 1X GlutaMax, 0.5X B-27 Minus Vitamin A, 0.5X N2, 0.1 mM 2-Mercaptoethanol, and 100 μg/ml Normocin (product numbers are listed in Table S2) [7]. The number of HF buds and the length of HF showed no significant difference between the two media under the same total number of cells (Fig. 2D and E). However, the area of pigmented hair bulbs was significantly larger in the OMM-cultured organoids compared to those in Advanced DMEM/F12 (Fig. 2F). This suggests that the addition of Neurobasal medium, traditionally used for culturing neural cells (Nestin-positive cells), might be a regulatory factor contributing to the enhanced pigmentation of HFs. This observation is particularly intriguing given the known multipotency of Nestin-positive cells, which are not only capable of neural differentiation but also have the potential to give rise to melanocytes, the pigment-producing cells [6]. Using Oil Red O staining, we observed the formation of cavities within the mHFO, along with pigmented HFs, sebaceous glands, sweat ducts and adipocytes on the surface (Fig. 2G). The expression of HFs keratin KRT5, the dermal marker vimentin, neuro-glial-associated markers p75, and the proliferating cell marker Ki67 were all present in the mHFO (Fig. 2H). Interestingly, mHFO expressed several specific markers of the arrector pili muscles (APM, α-SMA + , ITGA8 +) in HF and even exhibited distinct contraction phenomena (movie S1; Fig. S2A). Furthermore, these APM expressed the temperature-sensing receptor TRPV2 (Fig. S2B). The evidence above indicates that these organoids possess fully functional skin characteristics, providing a valuable model for the subsequent study of melanocyte development.
Fig. 1
Schematic representation of the preparation of mHFO. To create mHFO, isolated newborn mouse epidermal and dermal cells were combined 1:1 with 2% Matrigel, 1% GlutaMAX-1, and 0.2% Normocin, then cultivated in 96 U-bottom well plate
Fig. 2
mHFO culture and characterization. A Morphological changes during the life cycle of mHFO under the culture of Advanced DMEM/F12 (Adv). The hair follicle (HF) germinates in about a week, grows rapidly and vigorously in 1–2 weeks, and shrinks after 2 weeks. The bottom left corner is a magnified image of the HF in the rectangular box. B HF morphology of mHFO under Adv vs. OMM culture on day9. The bottom left corner is a magnified image of the HF in the rectangular box. C Macroscopic views of mHFO after incubation with Adv and OMM on day13. D The number of HF in a single mHFO under Adv and OMM culture conditions. HF were counted after 13 days of culture. For organoids generated with identical total cell numbers, there was no significant difference in the number of HFs between the Adv and OMM culture conditions (Student's t-test, ns: no significant difference, n = 3). Within each culture condition, organoids with total cell numbers of 20 × 10^3 and 40 × 10^3 exhibited a significantly higher number of HFs compared to those with 10 × 10^3 cells (Tukey’s test; **p < 0.01, ***p < 0.001, n = 3). E HF length was evaluated in the total number of 10 × 10^3 cell following a week of Adv and OMM culture. The calculation of HF lengths was obtained using image J (Student's t-test; ns: no significant difference, n = 16). F Hair bulb pigmentation area was evaluated in the total number of 10 × 10^3 cell following a week of Adv and OMM culture. The data was calculated using image J (Student's t-test; ***p < 0.001, n = 16). G The slices of mHFO were stained with oil red O and DAPI observed using a fluorescence microscope. The center of the mHFO forms a cell-free stratum corneum, while features such as adipocytes, sweat ducts, sebaceous gland, and HF were visible at the sphere's edge. H mHFO was sectioned and stained with KRT5, Vimentin, p75 and Ki67. The inserted images show magnified views of the box area in the images. Scale bars: 100 µm
BMP4 effect on mHFO morphology and melanin productionThe morphology of mHFO after five days of continuous induction with BMP4, FGF2, SB431542, and LDN is shown in Fig. 3A and B (product numbers are listed in Table S3). In the BMP4-treated group, there was an increase in pigmentation in the hair bulb region, accompanied by a decrease in the number of HF. The addition of LDN and SB431542 disrupted existing HF structures, with some mHFOs showing signs of cell death. In the FGF2-treated group, the original cell assembly pattern was altered, resulting in a reduced number of HFs and the appearance of numerous scattered fibroblasts, some of which exhibited vigorous contractions (Fig. 3 A and C; movie S1–S5). To quantify melanin content, we collected 24 mHFOs and analyzed them using the NaOH lysis method. BMP4 treatment significantly increased total melanin levels compared to the control group. This effect of BMP4 on melanin synthesis aligns with its established role as a key signaling molecule in melanocyte biology, where it regulates the expression of melanogenic enzymes and related transcription factors, such as MITF [10]. Conversely, LDN, a BMP4 inhibitor, reduced melanin content compared to the control group, while SB431542 and FGF2 had no significant effect on melanin levels (Fig. 3D). Additionally, the observed decrease in HF quantity and length suggests that the competitive balance between BMP and WNT signaling in the bulge region is critical for controlling stem cell homeostasis and cyclic activation (Fig. 3E and F) [11]. Notably, BMP4 also increased the pigmented area of the HFs and enlarged their size (Fig. 3G and H). These results indicate that BMP4 regulates HF growth by influencing their number, length, pigmentation, even the size.
Fig. 3
Effect of individual treatments with BMP4, FGF2, LDN, and SB431542 on HF pigmentation in cultured mHFOs (day15). A Representative images of mHFO after treatment with BMP4, FGF2, LDN, and SB431542 for 7 days, compared to control. The top row shows overall morphology, while the bottom row highlights pigmentation (red arrows indicate overpigmented areas). Scale bars: 100 µm. B Gross morphology of HF in treated organoids as seen in culture tubes under different conditions. C Histological sections of mHFO organoids stained with H&E. The lower panels provide magnified views of the boxed regions in the upper panels, highlighting HF structure. D Quantification of total melanin content 24 mHFO under different treatment conditions. (Dunnett’s test; *p < 0.05, **p < 0.01, ns: no significant difference, n = 3). E Number of HF formed in organoids treated with BMP4, showing a significant reduction with BMP4 treatment (Student's t-test; **p < 0.01, n = 8). F–H Length, area of pigmented hair bulbs, and size of HF relative to control after BMP4 treatment (Student's t-test; ***p < 0.001, n = 16). BMP4 treatment significantly reduced the length (F) while increasing the area of pigmented hair bulbs (G) and HF size (H)
BMP4-induced nestin changes in the HFNestin, an intermediate filament protein, is predominantly expressed in neural stem cells and is recognized as a key marker of multipotency, reflecting the cell's potential to differentiate into various lineages. In HF, Nestin-positive cells are localized within the bulge region and DP, and in sympathetic nerve fibers associated with the APM. These stem cells are integral not only to the intricate interactions with the peripheral nervous system but also to the regulation of the HF growth cycle. Notably, BMP4 is expressed in the DP and bulge regions, suggesting that the balance between stemness, indicated by Nestin expression, and differentiation towards a melanocyte lineage may be regulated by BMP4 (Fig. 4A–C).
Fig. 4
BMP4 modulates Nestin-Positive cell differentiation in mHFO. A Localization of Nestin-positive cells in HF structures. Nestin-positive cells (green) around the bulge (Bu), dermal papilla (DP), and sympathetic nervous system (SNS) near the arrector pili muscle (APM). Scale bars: 50 µm. B BMP4 (gray) co-localizes with Nestin (green) in HF. Scale bars: 100 µm. C Schematic representation of HF structure showing the localization of Nestin + cells (pink) and BMP4 + cells (green) in relation to the Bu, SNS, DP and hair shaft (HS). D Confocal microscopy images of mHFO derived from Nestin-Cre transgenic mice, cultured under normal conditions (Adv) on day 4. The images show the distribution of Nestin + (green) and tdtomato + (red) cells. Insets provide magnified views of the selected regions. The dashed line highlights the DP region. Scale bars: 100 µm. E, F Comparison between control and BMP4-treated mHFO shows altered Nestin (green) expression on day 6 (E) and day 9 (F). The upper panels show organoids cultured under normal conditions, while the lower panels show organoids treated with 40 ng/mL BMP4 for 2 days and 5 days. Bu outlined with dotted lines; DP with dashed lines. Inset shows magnified view. Scale bars: 100 µm. G Quantification of Nestin + cell fluorescence intensity in HF organoids on day 9 under control and BMP4-treated conditions. A significant decrease in Nestin + fluorescence intensity is observed in BMP4-treated samples (Student's t-test; ***p < 0.001, n = 10)
The above findings suggest that the addition of Neurobasal medium for neural cells (Nestin +) might be a regulatory factor in HF pigmentation. Moreover, BMP4 has been identified as a key cytokine in controlling HF pigmentation. Therefore, we hypothesize that BMP4 might control the commitment of melanocyte stem cells (MeSCs) within HF by influencing the differentiation of Nestin-positive stem cells. The distribution of Nestin-positive cells in the HF is consistent with that in the constructed mHFO (Fig. 4D–F). On day 4 of mHFO culture, HF buds emerge, with Nestin being enriched in the DP region. By day 6, Nestin appears in the bulge area. On day 9, as the HF elongates, Nestin distribution within the follicle becomes more distinct. Statistical analysis reveals that BMP4 treatment reduces the average fluorescence intensity of Nestin in the HF (Fig. 4G).
An increase in CD117 + melanocytes was observed over time, peaking on day 15 (Fig. 5A). mHFOs from day 6 was treated with BMP4 for one week, resulting in a significant decrease in Nestin content and a marked increase in the number of CD117 + melanocytes (Fig. 5B). Nestin-positive cells isolated from mice were subjected to in vitro induction. Under the intervention of BMP4, cells initially co-expressing DCT, a melanocyte marker, and Nestin gradually expressed only DCT. This suggests that BMP4 may guide the differentiation of these cells, directing them toward melanocytes (Fig. 5C and D).
Fig. 5
BMP4-induced differentiation of Nestin-positive into melanocytes in mHFOs and isolated cells. A Flow cytometry plots showing the percentage of CD117-positive melanocytes in organoids at days 6, 9, and 15. The bar graph quantifies the increase in CD117 + melanocytes over time, with a significant expansion by day 15 (Tukey’s test; ***p < 0.001, n = 3). B Comparative of Nestin-positive cells and CD117-positive melanocytes of mHFO after BMP4-treated 6 days. BMP4 treatment significantly decreases the proportion of Nestin + cells and increases CD117 + melanocytes (Student's t-test; *p < 0.001, **p < 0.0001, n = 7). C Immunofluorescence images of untreated cells stained for DAPI (blue), DCT (red), and Nestin (white). Co-localization of DCT and Nestin indicating the presence of undifferentiated cells. Scar bars: 20 μm. D Immunofluorescence images of BMP4-treated cells. It highlights the differentiation induced by BMP4 treatment, with a shift towards melanocyte lineage. Scar bars: 20 μm
Bulge nestin-positive cells as the melanocyte sourceCD117 is crucial for melanocyte development, contributing to melanocyte migration, survival, proliferation, and differentiation. In the HF, CD117 is expressed in key regions: the sympathetic nerves associated with the APM, the nerve plexus connected to the DP, and notably, the bulge region, which acts as a stem cell niche and reservoir for MeSCs. Similarly, Nestin is expressed in the sympathetic nerves, bulge, DP, and the nerve plexus near the DP (Fig. 6A and B). The bulge is a focal area of co-expression for CD117, Nestin, and DCT, emphasizing its role as a significant source of melanocytes within the HF (Fig. 6B). This co-localization underscores the bulge’s importance as a niche that preserves melanocyte progenitors and supports their differentiation as needed. A schematic diagram that visually represents the spatial distribution of CD117 + cells, Nestin + cells, and DCT + cells within the HF, illustrates their respective locations and interactions within this niche (Fig. 6C). This configuration of cells and markers reinforces the bulge’s function as a key site for melanocyte regeneration and supports the dynamics of HF stem cell regulation.
Fig. 6
Nestin-positive cells in the bulge region as a potential source of melanocytes. A Immunofluorescence staining of HF from Nestin-Cre transgenic mice showing the distribution of Nestin + (green) and CD117 + (white) cells. The upper panels focus on the SNS, while the lower panels highlight the DP. DAPI (blue) marks the nuclei. Arrowheads indicate SNS or DP. Scale bars: 50 µm. B Immunofluorescence analysis showing the spatial relationship between Nestin+, CD117+, and Nestin+ cells in the HF of Nestin-Cre transgenic mice. High-magnification images of the bulge (Bu) region in HF, illustrating the co-localization of Nestin + (green) and CD117 + (white) cells. The red downward arrow indicates the migration direction of DCT⁺ melanocyte stem cells from the bulge toward the hair germ. Arrowheads indicate Bu region. HG, Hair grem. Scale bar: 50 µm. C Schematic representation of HF structure in Nestin-Cre transgenic mice, depicting the localization of DCT+ cells (red), CD117 + cells (purple) and Nestin + cells (green) in relation to the Bu, DP, and SNS
留言 (0)