
記住我
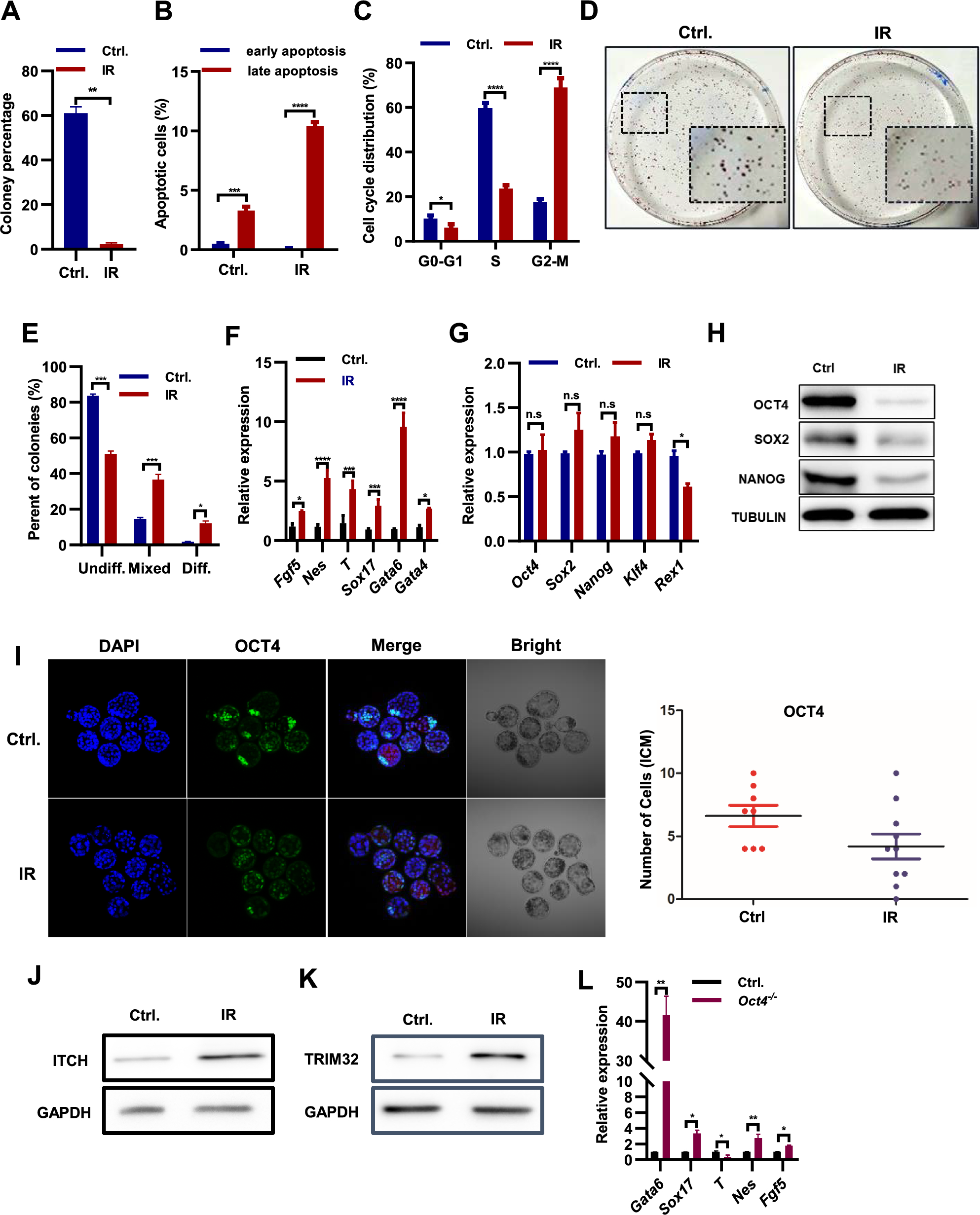


To evaluate the impact of X-ray irradiation on the self-renewal capacity, ESCs were first exposed to irradiation and colony formation monitored, decreased numbers of ESC colonies indicated self-renewal ability was significantly impaired (Fig. 1A; Suppl. Fig. S1A). This decline in colony formation coincided with an elevated incidence of apoptosis, as the percentage of apoptotic cells was found to be higher in irradiated ESCs (~ 10.4%) compared to non-irradiated cells (~ 3.2%) (Fig. 1B). Furthermore, the irradiation treatment induced alterations in the cell cycle, characterized by a 51.19% increase in the proportion of cells in the G2-M cell cycle phases and a decrease of 36.12% in the S phase (Fig. 1C). In addition to the diminished colony-forming potential, alkaline phosphatase (AP) staining revealed a reduction in the proportion of homogeneously stained, undifferentiated colonies in irradiated ESCs (Fig. 1D–E). Collectively, our findings underscore the detrimental effects of X-ray irradiation on ESC self-renewal, as evidenced by increased apoptosis, perturbations in the cell cycle, and compromised colony-forming ability.
Fig. 1
Irradiation-induced impairment of ESC self-renewal and promotion of ESC differentiation. A percentage of survived ESC colony formed upon 4 Gy x-ray irradiation treatment. Certain number of ESCs were irradiated with 4 Gy x-ray, and then plated for colony formation assay. B Percentage of ESCs at early and later apoptosis stages after irradiated with 4 Gy x-ray. C cell cycle assay of ESCs upon 4 Gy x-ray irradiation treatment. D AP staining of irradiated ESCs. The ESCs were irradiated with a dose of 4 Gy of x-rays before being plated for the single colony assay. E Percentage of undifferentiated, partially differentiated and fully differentiated colony formed of ESCs upon irradiation with 4 Gy x-ray. F–G qPCR analysis of indicated lineage marker genes (F) and pluripotency genes (G) in ESCs upon irradiated with 8 Gy x-ray twice in 48 h. H Western blot analysis of indicated proteins in ESCs upon irradiated with 8 Gy x-ray twice in 48 h. I Immunostaining analysis of E4.5 embryos with OCT4 antibody following dual 2 Gy x-ray irradiation at E2.5 and E3.5. Scale bar: 50 µm. J–K Western blot analysis of ITCH (J) and TRIM32 (K) in ESCs upon irradiated with 8 Gy x-ray twice in 48 h. L qPCR analysis of indicated lineage marker genes in control (WT) and Oct4−/− ESCs
Irradiation-induced upregulation of Itch and Trim32 promotes the differentiation of ESCsThe observed decrease in the percentage of undifferentiated colonies in irradiated ESCs serves as evidence that irradiation treatment stimulates ESC differentiation (Fig. 1E). To investigate the effect of irradiation on ESC differentiation, we assessed the expression levels of established lineage markers. The transcript levels of the endoderm markers Gata4, Gata6, and Sox17, the mesoderm marker Brachyury (T), and the ectoderm markers Fgf5 and Nes were notably upregulated in ESCs following exposure to X-ray irradiation at the specified dosages when compared to their corresponding control groups (Fig. 1F; Supple. Fig. S1B). These findings demonstrate that irradiation promotes ESC differentiation across all three lineages. Consistent with this, the expression of Rex1, a well-known marker gene for the undifferentiated state of ESCs, notably decreased in irradiated ESCs (Fig. 1G). However, qPCR analysis revealed minimal effects of irradiation on the transcript levels of core pluripotency genes Oct4, Sox2, Nanog, and Klf4 (Fig. 1G). Therefore, we examined the protein levels of OCT4, SOX2, and NANOG. In line with the induced expression of differentiation marker genes in ESCs following X-ray irradiation treatment (Fig. 1F), the protein levels of OCT4, SOX2, and NANOG significantly decreased in irradiated ESCs (Fig. 1H). Furthermore, irradiated E4.5 mouse embryos exhibited reduced OCT4 and NANOG positive cells (Fig. 1I; Supple. Fig. S1C, D). These findings indicate that irradiation promotes ESC differentiation by modulating the stability of key pluripotency proteins.
As an inhibitor of 26S proteasome, MG132 reduces the degradation of ubiquitin-conjugated proteins in ESCs [19]. The addition of MG132 reduced the degradation of OCT4, NANOG and SOX2 (Supple. Fig. S1E), indicating the induced ubiquitination of key pluripotency proteins. WWP2, ITCH, and TRIM32 are E3 ubiquitin ligases known to facilitate the degradation of OCT4 protein [20]. To investigate whether irradiation promotes OCT4 degradation by modulating the expression of Wwp2, Itch, and Trim32, we examined their expression levels in irradiated ESCs. Following irradiation, we observed upregulation of both transcript and protein levels of Itch and Trim32 genes (Supple. Figs. S1F, 1J, K), which was further confirmed by immunostaining experiments (Supple. Fig. S1G, H). However, the transcript level of the Wwp2 gene did not exhibit significant changes (Supple. Fig. S1F). Deletion of the Oct4 gene induced the expression of Gata6, Sox17, FGF5, and Nes in ZHBTc4 ESCs upon Doxycycline treatment (Fig. 1L) [21], suggesting that irradiation-induced ESC differentiation may occur through downregulation of the OCT4 protein. However, the expression of the mesoderm marker gene T, which increased following irradiation treatment (Fig. 1F), was downregulated upon Oct4 gene knockout (Fig. 1L), indicating the involvement of additional mechanisms by which irradiation treatment induces ESC differentiation. In summary, irradiation exerts a promotional effect on the differentiation of embryonic stem cells through the modulation of E3 ubiquitin ligase activity, ultimately resulting in the degradation of pluripotency proteins.
Irradiation promotes the differentiation of ESCs via upregulating the expression of Yap1To elucidate the underlying mechanism by which irradiation regulates ESC differentiation, we performed RNA sequencing (RNA-seq) to examine global gene expression changes in irradiated ESCs. Our analysis revealed 2627 significantly downregulated genes and 2265 upregulated genes in irradiated ESCs compared to control ESCs (Fig. 2A; Supplementary Table 1). Consistent with the observed induction of apoptosis following irradiation treatment (Fig. 1B), Gene Ontology (GO) analysis demonstrated enrichment of cell death among upregulated genes in irradiated ESCs (Fig. 2B upper panel). Upregulated genes were found to be associated with cell differentiation and cell proliferation (Fig. 2B, upper panel), indicative of an increased propensity for ESC differentiation and alterations in the cell cycle dynamics (Fig. 1C–F). Down-regulated genes were associated with apoptotic signaling pathway (Fig. 2B, lower panel). The expression of genes associated with the Hippo pathway in ESCs was observed to increase under irradiation treatment (Fig. 2A), suggesting a crucial role of this pathway in promoting ESC differentiation under such conditions. Correspondingly, both mRNA and protein levels of Yap1, a transcriptional co-activator of the Hippo pathway [22], were significantly upregulated in irradiated ESCs (Fig. 2C; Supple. Fig. S2A). This upregulation of Yap1 resulted in an enhanced binding of YAP1 to its target genes Ankrd, Axl, Ptx3, Cyr61, and Ctgf (Fig. 2D), thereby leading to their upregulation in irradiated ESCs. Previous studies have demonstrated that overexpression of Yap1 in ESCs disrupts their self-renewal and triggers differentiation by upregulating the expression of lineage-specific genes [22, 23]. Hence, we hypothesized that the regulation of Yap1 expression might be involved in the irradiation-induced changes in the differentiation potential of ESCs. To test this hypothesis, we generated mouse ES cell lines with ectopic expression of either human or mouse Yap1. As expected, we observed an upregulation of the YAP1 target genes in these cells (Supple. Fig. S2B, C). Additionally, in line with a previous report [22], ectopic expression of both mouse and human Yap1 led to differentiation morphological changes in ESCs (Supple. Fig. S2D, E). The expression of core pluripotency regulators, such as Sox2, Nanog, Tbx3, Klf4, Klf5, and Klf2, significantly decreased in both human and mouse Yap1-overexpressing ESCs (Supple. Fig. S2F, G), indicating that ectopic expression of Yap1 impairs self-renewal and promotes ESC differentiation. Consistent with these findings, AP staining revealed a decrease in homogeneously stained, undifferentiated colonies in human YAP1-overexpressing ESCs (Supple. Fig. S2H). Furthermore, both human and mouse Yap1-overexpressing ESCs showed increased expression of established lineage marker genes (Fig. 2E; Supple. Fig. S2I). Interestingly, Yap1-overexpressing ESCs subjected to irradiation treatment exhibited a further decrease in the protein level of pluripotency factors OCT4, SOX2, and NANOG compared to wild-type ESCs (Supple. Fig. S2J). Additionally, the expression of lineage marker genes Fgf5, Nes, T, Sox17, and Gata6 was significantly higher in irradiated ESCs with human or mouse Yap1-overexpression than in wild-type ESCs and irradiated wild-type ESCs (Fig. 2F (human); Supple. Fig. S2K (mouse)). Collectively, these findings suggest that irradiation treatment upregulates Yap1 expression, leading to impaired self-renewal and increased differentiation of ESCs.
Fig. 2
Irradiation enhances ESC differentiation through upregulation of Hippo signaling pathway. A Heat map depicts the changes in gene expression in mESCs irradiated with 8 Gy x-ray twice in 48 h. The color represents the Z score (row-wise) of the log2 FPKM values of the 4892 genes affected. B GO ontology analysis for biological processes associated with genes whose expression changed in ESCs upon irradiation treatment in A. C qPCR analysis of transcript levels of Yap1 and its target genes in ESCs irradiated with 8 Gy twice in 48 h. D YAP1 levels at indicated target genes in ESCs following dual 8 Gy x-ray irradiation within 48 h determined by ChIP-qPCR. E qPCR analysis of transcript levels of lineage marker genes in ESCs and ESCs overexpressing human YAP1 gene (ov-hYAP1 ESCs). F qPCR analysis of transcript levels of lineage marker genes in ESCs with or without overexpressing human YAP1 gene following dual 8 Gy x-ray irradiation within 48 h determined by ChIP-qPCR. G Binding distribution of YAP1 on the genome based on its ChIP-seq analysis. H–I Genome browser view of YAP1 and H3K4me3 at the Fgf5 (H) and Gata6 (I) loci in ESCs; (J) YAP1 levels at indicated target genes in ESCs following dual 8 Gy x-ray irradiation within 48 h determined by ChIP-qPCR
To investigate the underlying molecular mechanisms of Yap1-mediated ESC differentiation, we analyzed YAP1 chromatin immunoprecipitation sequencing (ChIP-seq) data and found that 41% of YAP binding occurred within the promoter regions of genes (Fig. 2G), indicating that Yap1 primarily regulates gene expression by controlling their promoter activities. Further analysis revealed that YAP1 binds to the promoter regions of lineage marker genes Fgf5, Gata6, T, and Nes in ESCs (Fig. 2H, I; Supple. Fig. S2L–M). ChIP-qPCR experiments revealed that irradiation treatment resulted in an increased YAP binding on the promoters of the lineage markers Fgf5, Nes, T, Mixl1, Sox17, and Gata6 (Fig. 2J), suggesting that irradiation treatment may increase Yap1 expression and upregulate the expression of lineage marker genes through enhanced binding of YAP1 to their promoter regions.
Irradiation lifts the inhibition of BAF complex on Yap and its target genes in mESCsThe BAF complex factor ARID1A has been shown to bind to and inhibit the oncogenic transcriptional coactivators YAP and TAZ in cancer cells [24]. We speculated whether BAF complex inhibits the transcription of Yap1 in ESCs. Co-immunoprecipitation (Co-IP) analysis demonstrated the interaction of YAP1 with BAF complex components BRG1 and DPF2 in mESCs (Fig. 3A). Deletion of Brg1, Dpf2, or Arid1a in ESCs resulted in increased expression of Yap1 and its target genes Ankrd, Ptx3, Cyr61, and Ctgf (Fig. 3B; Supple. Fig. S3A, B), indicating that the BAF complex inhibits the expression of Yap1 and its target genes in mESCs. We hypothesized that the BAF complex may prevent the binding of YAP1 to its target genes. To test this idea, we analyzed the ChIP-seq data of BRG1, DPF2, and YAP1 and identified 1,064 co-binding genomic locations (Fig. 3C, D), which included typical Yap1 target genes such as Cyr61 and CTGF (Fig. 3E). The binding of YAP1 to its target genes increased in Dpf2 and Brg1 knockout ESCs (Fig. 3F; Supple. Fig. S3C). Therefore, the BAF complex represses the expression of Yap1 target genes by preventing the binding of YAP1 to them.
Fig. 3
Irradiation exerts control over Yap target genes by modulating the binding of the BAF complex to them. A Co-immunoprecipitation with BRG1 antibody revealed its interaction with DPF2 and YAP1. B qPCR analysis of Yap1 and its target genes in WT and Brg1−/− ESCs. C Venn diagram depicting the number of peaks that are bound by YAP1, BRG1 and DPF2 from their ChIP-seq analyses. D Heat map of the ChIP-seq signals of BRG1, DPF2 and YAP1 around their common binding sites. The color represents the log2 RPM values. E Genome browser view of YAP1, BRG1 and DPF2 at the Cyr61 and CTGF loci in ESCs. F YAP1 levels at indicated target genes in WT and Brg1.−/− ESCs determined by ChIP-qPCR. G BRG1 levels at indicated target genes in ESCs following dual 8 Gy x-ray irradiation within 48 h determined by ChIP-qPCR. H ESCs were transfected with I- Sce I plasmid. ChIP assay was conducted 5 and 8 h after I-Sce I transfection, and qPCR analyses were used to detect the enrichment of BRG1 relative to the IgG control. I qPCR analysis of YAP1 target genes in ESCs (Ctrl.), irradiated ESCs (Ctrl.IR), and irradiated ESC overexpressing Yap1 (ov-Yap1, IR). J qPCR analysis of Yap1 target genes in ESCs (Ctrl.), irradiated WT ESCs (Ctrl. IR) and Brg1 KO ESCs (Brg1 KO IR)
Irradiation treatment upregulates the expression of Yap1 and its target genes by increasing Yap1 expression (Fig. 2C, D), the molecular mechanism of which remains unknown. Considering the inhibitory role of the BAF complex in the binding of YAP1 to its targets and the regulation of their expression (Fig. 3F; Supple. Fig. S3C), we speculated that irradiation treatment may either decrease the expression of BAF complex components or alleviate the binding of the BAF complex to YAP1 target genes, thereby increasing the binding of YAP1 to its targets. Irradiation did not alter the expression of Brg1, Dpf2, and Arid1a at both mRNA and protein levels (Supple. Fig. S3D, E). Indeed, irradiation treatment reduced the binding of BRG1 to YAP1 and its target genes (Fig. 3G); concurrently, the binding of YAP1 to those locations increased (Fig. 2D). In conclusion, irradiation upregulated the expression of YAP1 target genes by reducing the binding of the BAF complex and increasing the binding of YAP1 to these targets.
Multiple studies have indicated that BAF complexes play crucial roles in facilitating the efficacy of the DNA damage response [25]. The recruitment of ARID1A to DNA double-strand breaks (DSBs) occurs through its interaction with the upstream DNA damage checkpoint kinase ATR [26]. We hypothesized that irradiation-induced DNA damage may recruit the BAF complex to DNA breaks from its binding sites associated with Yap1 and YAP1 target genes, thereby alleviating its inhibition on YAP1 binding. To investigate this, we employed a ChIP assay to examine whether BRG1 was recruited in the vicinity of a site-specific I-SceI-induced DSB [26]. The DR-GFP construct, containing a cutting site for the I-SceI restriction enzyme, was stably integrated into ESCs (Supple. Fig. S3F). ChIP-qPCR analysis revealed enrichment of BRG1 following I-SceI transfection (Fig. 3H). Thus, irradiation-induced DNA damage may recruit BRG1 away from its binding sites on Yap1 and its target genes, leading to increased binding of YAP1 and upregulation of Yap1 and its target genes. Additionally, compared to wild-type ESCs, irradiation treatment of Yap1-overexpressing ESCs further enhanced the expression of Yap1 target genes (Fig. 3I), supporting the notion that IR treatment disrupts the binding of the BAF complex and increases the binding of YAP1 to Yap1 target genes. Furthermore, consistent with these findings, irradiation treatment of Brg1, Dpf2, and Arid1a knockout ESCs resulted in further elevation of the expression of Yap1 target genes, including Ankrd, Ptx3, Cyr61, and Ctgf (Fig. 3J; Supple. Fig. S3G, H), providing further evidence that the deletion of BAF subunits enhances the accessibility of Yap1 to its targets. Therefore, we conclude that irradiation regulates the expression of Yap1 and its target genes by modulating the binding of the BAF complex to these sites.
Irradiation promotes the differentiation ESC via activating Wnt signaling pathwayFurthermore, GO analysis of the upregulated genes showed enrichment for the Wnt signaling pathway (Fig. 2B). This observation was corroborated by qPCR analysis, which revealed elevated expression levels of Wnt3, Wnt3a, and several Wnt target genes, including Cdx1, Sp5, Msx2, Axin2, Cdh2, Lrp2, and Ctnb1, in irradiated ESCs (Fig. 4A, B). Consistently, the protein level of β-CATENIN was increased in irradiated ESCs (Fig. 4C). Collectively, these findings indicate that irradiation treatment enhances the activity of Wnt pathway in ESCs.
Fig. 4
Irradiation treatment induces increased activity of Wnt and TGF-β signaling pathways in ESCs. A qPCR analysis of the expression levels of Wnt3 and Wnt3a transcripts in ESCs (Ctrl.), and ESCs following irradiation with 8 Gy x-ray once in 24 h and twice in 48 h (IR). B qPCR analysis of indicated Wnt target genes in ESCs irradiated with 8 Gy x-ray twice in 48 h. C Western blot revealed the protein levels of β-CATENIN in ESCs (Ctrl.) and irradiated ESCs (IR). D qPCR analysis of the typical lineage genes in ESCs (Ctrl.), irradiated ESCs (IR), and irradiated ESCs with the addition 10 μg/ml of IWP2 (IR + IWP2). E qPCR analysis of transcript levels of the typical lineage genes in WT ESCs (Ctrl.), irradiated WT ESCs (IR), and irradiated ESCs overexpressing Wnt3a (IR, ov-Wnt3a). F qPCR analysis of transcript levels of Wnt target genes in WT (Ctrl.) and Dpf2 KO ESCs. G BRG1 levels at indicated Wnt target genes in WT (Ctrl.) and irradiated ESCs (IR) determined by ChIP-qPCR. H qPCR analysis of transcript levels of Wnt target genes in WT (Ctrl.), irradiated WT (IR) and irradiated Dpf2 KO ESCs (Dpf2.−/− IR). I BRG1 levels at indicated lineage specific genes in irradiated ESCs determined by ChIP-qPCR. J–K Genome browser view of YAP1, β-CATENIN, BRG1 and DPF2 at the T (J) and Fgf5 (K) loci in ESCs. L Co-immunoprecipitation with BRG1 antibody revealed its interaction with β-CATENIN and YAP1. M qPCR analysis of transcript levels of Tgfb2, Tgfb3, Inhba and Nodal genes in WT (Ctrl.), and WT ESCs irradiated with x-ray treatment (IR). N qPCR analysis of transcript levels of lineage marker genes in WT (Ctrl.), irradiated WT ESCs (IR), and irradiated WT ESCs in the presence of the TGF-β receptor inhibitor SB431542 at a concentration of 10 μM (IR + SB431542)
The knockdown of OCT4 in human ESCs has been shown to activate the β-catenin signaling pathway and facilitate ESC differentiation [27]. Conversely, inhibition of the Wnt signaling pathway prevents the activation of mesendoderm differentiation [28]. Based on these findings, we hypothesized that increased Wnt signaling may promote ESC differentiation under irradiation treatment. Indeed, we observed enhanced binding of β-CATENIN to typical lineage marker genes in irradiated ESCs (Supple. Fig. S4A). Furthermore, the addition of the Wnt signaling inhibitor IWP2 during irradiation treatment reduced the expression of lineage marker genes Nes, Fgf5, Mixl1, Gata6, and Sox17 (Fig. 4D), highlighting the crucial role of Wnt signaling in regulating ESC differentiation under irradiation treatment. Consistently, overexpression of Wnt3a further increased the expression of lineage markers such as Gata4, Gata6, Sox17, T, and Nes in ESCs upon irradiation treatment (Fig. 4E). Overexpression of Wnt3 increased the expression of Gata6 and Sox17, but not the mesoderm gene T and ectoderm gene Fgf5 under irradiation treatment conditions (Supple. Fig. S4B). The Wnt signaling pathway has previously been reported to maintain ESC self-renewal [29]. Overexpression of Wnt3a repressed the expression of endoderm marker genes Gata4, Gata6, and Sox17, as well as the mesoderm gene T, in ESCs without irradiation treatment (Supple. Fig. S4C). This suggests that Wnt3a promotes endoderm differentiation of ESCs only after their differentiation has been initiated. To further investigate, we cultured doxycycline-inducible Wnt3a ESCs in non-LIF medium for 24 h to initiate differentiation and then induced Wnt3a overexpression upon the addition of doxycycline for 48 h. qPCR analysis revealed that overexpression of Wnt3a increased the expression of endoderm marker genes Gata4 and Gata6, as well as the mesoderm marker genes T and Mixl1, in differentiating ESCs, while the expression of the neural ectoderm marker gene Nes and the ectoderm gene Fgf5 remained unchanged (Supple. Fig. S4D). Similarly, when inducing EB formation with doxycycline-inducible Wnt3a ESCs in non-LIF medium for 24 h before inducing Wnt3a overexpression for 48 h, qPCR analysis showed that overexpression of Wnt3a increased the expression of endoderm marker genes Gata4 and Gata6, as well as the mesoderm marker gene T, in 3-day EBs, while the expression of the neural ectoderm marker gene Nes decreased and the ectoderm gene Fgf5 did not change (Supple. Fig. S4E). Thus, we conclude that irradiation promotes ESC differentiation by activating the Wnt signaling pathway.
The expression of Wnt3 and Wnt3a, as well as the target genes associated with the Wnt pathway, were found to be upregulated in ESCs following irradiation treatment (Fig. 4A, B). Furthermore, the deletion of Dpf2 or Brg1 in ESCs resulted in an increase in the expression of Wnt pathway target genes (Fig. 4F; Supple. Fig. S4F). ChIP-qPCR analysis using an antibody against BRG1 demonstrated a decrease in the binding of BRG1 to Wnt target genes upon irradiation treatment (Fig. 4G). IR treatment of Dpf2 knockout ESCs resulted in further elevation of the expression of Wnt target genes, including Axin2, Cdh2, Msx2, Sp5, Ctnnb1, and Lrp2 (Fig. 4H), providing further evidence that the deletion of BAF subunits enhances the accessibility of β-CATENIN to its targets. These findings indicate that, similar to the regulation of Yap1 and its target genes, the BAF complex represses the expression of Wnt target genes.
Both the Yap and Wnt pathways play crucial roles in regulating ESC differentiation in response to irradiation treatment (Figs. 2E, F, 4D, E). Following irradiation, there is an increase in the binding of YAP1 and β-CATENIN to typical lineage genes (Fig. 2J; Supple. Fig. S4A), while the binding of BRG1 decreases (Fig. 4I). Notably, the components of the BAF complex, BRG1, and DPF2, co-bind with YAP1 and β-CATENIN at lineage marker genes such as Sox17, T, Fgf5, and Nes (Fig. 4J, K; Supple. Fig. S4G, H). Co-immunoprecipitation experiments using a BRG1 antibody further confirm its interaction with both YAP1 and β-CATENIN (Fig. 4L). Based on these collective findings, we can conclude that irradiation treatment effectively regulates ESC differentiation through several mechanisms. It enhances the binding of YAP1 and β-CATENIN to characteristic lineage marker genes, leading to an increase in their expression. Additionally, irradiation treatment releases the inhibition of the BAF complex, further contributing to the upregulation of gene expression.
Irradiation promotes the differentiation of ESCs via activating TGF-β signaling pathwayThe expression of genes associated with the TGF-β pathway in ESCs was observed to increase under irradiation treatment (Fig. 2B), suggesting a crucial role of this pathway in promoting ESC differentiation under such conditions. Correspondingly, the transcript levels of TGF-β pathway associated genes Tgfb2, Tgfb3, Inhba and Nodal were significantly upregulated in irradiated ESCs (Fig. 4M). Irradiation promoted the expression of typical marker genes of all three lineages Gata4, Sox17, T, Nes and Fgf5, which was restored upon the addition of SB431542, the inhibitor of TGF-β pathway (Fig. 4N), suggesting that irradiation promotes the differentiation of ESCs via activating TGF-β signaling pathway. Consistently, ChIP-seq analysis demonstrates the co-localization of SMAD2 with YAP1, β-CATENIN, BRG1 and DPF2 to lineage marker genes of three germ layers T, Fgf5, Sox17 and Nes (Fig. 4J–K; Supple. Fig. S4G, H). Co-IP experiments shown the interaction of SMAD2 with BRG1, YAP1 and β-CATENIN, suggesting the coordination of TGF-β, Hippo and Wnt pathways on the regulation of ESC differentiation. In line with this, the inhibition of TGF-β with SB431542 led to the restoration of irradiation-induced β-CATENIN enrichments at Sox17, Gata6, T, Fgf5, and Nes (Supple. Fig. S4I). Moreover, the elimination of Brg1 resulted in an increase in the expression of Tgfb2, Tgfb3, and Inhba (Supple. Fig. S4J), indicating the suppressive role of the BAF complex in the TGF-β signaling pathway in ESCs. Consistently, the upregulated expression of genes related to the TGF-β pathway such as Tgfb2, Tgfb3, Inhba and Nodal was observed in irradiated ESCs (Fig. 4M), accompanied by a significant reduction in the binding of BRG1 to these genes (Supple. Fig. S4K). Overall, irradiation-induced the alleviation of BAF inhibition on the TGF-β, Hippo, and Wnt signaling pathways, leading to their increased activity. The complex composed of the effector proteins from these three pathways collaboratively regulates the expression of characteristic lineage-specific genes.
Irradiation-induced expression of p53, and p63/p73 drives meso- and endo-dermal differentiation of ESCs, respectivelyUpon DNA damage, p53, p63 and p73 induce apoptosis and alter the distribution of the cell cycle, leading to a delay in cell growth [30]. In ESCs, irradiation upregulates both the transcript and protein levels of p53, p63, and p73 (Fig. 5A; Supple. Fig. S5A), which in turn may induce apoptosis and perturb the cell cycle (Figs. 1B, C, 2B). Previous studies have identified the Wnt signaling pathway as a target of p53 in mouse ESCs [28, 31]. Furthermore, Wnt-mediated Yap/β-Catenin transcription of Oct4 in ESCs is suppressed by the core hippo scaffold RASSF1A, itself a target of ATM in response to ionizing radiation [32], and switch YAP to support p73 mediated differentiation [23]. Based on this knowledge, we hypothesized that the upregulation of p53, p63, and p73 upon irradiation treatment might similarly switch Wnt signaling pathway away from pluripotency genes and towards differentiation, which was supported by the increased binding of β-CATENIN at lineage marker genes upon the IR treatment (Supple. Fig. S4A). Supporting this notion, overexpression of p53, p63, and p73 increased the transcript levels of Wnt3 and Wnt3a and the protein level of β-CATENIN (Fig. 5B; Supple. Fig. S5B). Irradiation treatment also elevated the expression of Itch and Trim32, leading to increased degradation of OCT4 protein in ESCs (Fig. 1H, J, K; Supple. Fig. S1E). Overexpression of p53, p63, and p73 further upregulated the expression of Itch, Trim32, and Wwp2, and the protein level of ITCH (Fig.
留言 (0)