
記住我
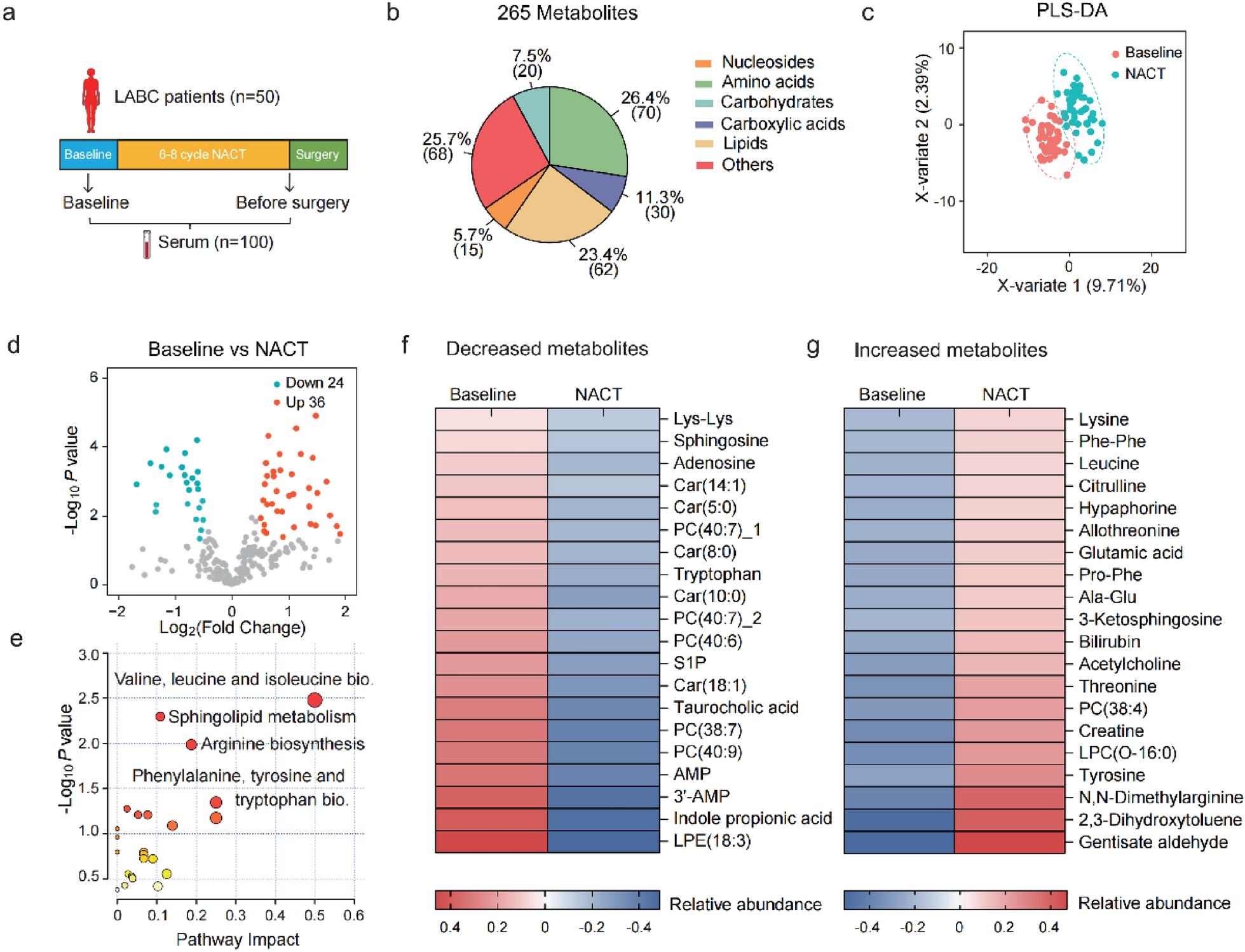



To study and compare sequential events during the early stage of GI and AI, we used a mouse model where wild-type females of FVB/N background at 8 weeks of age were set up for breeding. Following partum, litter size was standardized to 6 per dam [13]. Uniparous dams were randomly assigned to GI and AI cohorts. All 6 pups were removed from the AI dams on day 7 PPM. For GI dams, 3 pups each were removed on days 28 and 31 PPM, allowing for prolonged lactation. Multiple studies in the past analyzed early events (0–72 h post-involution) of involution specifically in the AI setting [18,19,20]. This study focused on mammary glands harvested intermittently between days 8.5 and 28 PPM from mice in both AI and GI cohorts (Fig. 1A). Lactating glands harvested on day 7 PPM (before removal of pups) were used as a baseline. Histological features of sequential stages of mammary gland involution undergoing AI vs. GI are captured in Fig. 1B.
Fig. 1
Histological analysis of mammary glands undergoing gradual vs. abrupt involution. A. Schematic diagram showing the workflow of mice subjected to abrupt involution (AI) and gradual involution (GI) followed by different harvest times. B. Representative images of H&E-stained sections of mouse mammary glands subjected to AI or GI and harvested at different days postpartum (PPM) as indicated. Scale bar = 200 μm C. Higher magnification images of H&E-stained section of GI mammary gland on indicated postpartum days. Day 8.5, 11 and 12: black arrows indicate large acini, and clear arrows indicate epithelial cells. Day17: clear arrows indicate epithelial cells, curved black arrows indicate a small group of adipocytes; Day 22: clear arrows indicate flattened epithelial cells, black arrows indicate the nuclear debris and arrowheads indicate repopulating adipocytes. Day 25: arrow heads indicate repopulating adipocytes; black arrows indicate immune cells. D. H&E-stained section of AI mammary gland on day 8.5: Grey arrow indicate lactating acini, black arrowheads indicate flattened epithelial cells and black arrows indicate karyorrhectic debris. Day 11: Grey arrows indicate collapsed alveoli, black arrows indicate immune cells and arrowheads indicate adipocytes. Day 12: arrowheads indicate adipose tissues, and black arrows indicate immune cells. Day17: arrow heads indicate adipose tissues. (C&D: Scale bar = 100 μm). E. Adipocyte repopulation over time, from day 7 to day 35 PPM. n = 6 per group per time point. F. Comparison of total number of adipocytes in AI and GI glands harvested on day 28 PPM. G. Comparison of total number of adipocytes categorized by size in AI vs.GI glands
Analysis of H&E-stained sections revealed that between days 8.5 and 12 PPM, GI glands were fully lactating and composed of numerous, densely packed, large acini lined by tall cuboidal epithelium and filled with amphophilic secretions, with almost no intervening adipose tissue (Fig. 1B and C). On day 17 PPM, the epithelium lining acini appeared less tall multifocally, and small groups of adipocytes were often present between acini (Fig. 1C). On day 22 PPM, acini were lined by low cuboidal to flattened epithelium, and nuclear debris became apparent in acinar lumens in the form of apoptotic bodies, suggesting acinar breakdown (Fig. 1C). Larger groups of adipocytes were present between acini (Fig. 1C, adipocytes quantified in Fig. 1E). By day 25, there was a significant increase in adipocyte repopulation, and few intact acini remained (Fig. 1C). In addition, a mixed inflammatory cell population was present multifocally surrounding partly incomplete acinar profiles and admixed with proteinaceous fluid and fat globules (Fig. 1C).
AI glands on day 8.5 PPM (1.5 days after pup removal) were also fully lactating with little intervening adipose tissue. However, acini were lined by flattened epithelium and there was abundant cytoplasmic and karyorrhectic debris admixed with few inflammatory cells indicating prominent acinar breakdown (Fig. 1B and D). On day 11 PPM, the glandular remodeling was more advanced with rare intact acinar profiles, increased numbers of inflammatory cells, and more abundant intervening adipose tissue (Fig. 1B and D). On day 12 PPM, there was a striking reduction of glandular parenchyma, with a significant increase in the area occupied by adipose tissue. Acinar breakdown and inflammatory cells were less prominent than on day 11 (Fig. 1B and D). These changes resembled those observed in GI-day 25 glands. On day 17 PPM, adipose tissue predominated largely in the AI glands and only ducts with a few acini were visible (Fig. 1B and D). The AI glands continued to remodel beyond day 17 and histologically resembled near a pre-pregnant state by day 28 PPM. On the contrary, the GI glands remodeled through day 31 and attained near pre-pregnant histology by day 35 PPM (Supplementary Fig. S1A). Signs of proliferative changes are apparent in day 31 and day35 AI glands that led to hyperplasia as reported earlier [13].
Quantification of adipocyte repopulation showed that in AI glands, repopulation spans over 10-days (day 12 to day 22 PPM), with a sharp increase in adipocyte repopulation in GI glands starting on day 25 and reaching a plateau on day 28 PPM, indicating complete remodeling of the gland (Fig. 1E). This repopulation was associated with simultaneous loss of mammary epithelial cells in both glands (Supplementary Fig. S1B). AI-day 28 glands exhibited a small but significant increase in adipocyte number compared to GI-day 28 glands (Fig. 1F). However, when categorized by size, there was no significant difference in the distribution of adipocytes between AI and GI glands on day 28 PPM (Fig. 1G), along with total adipocyte area, average size, and the number of crown-like structures (Supplementary Figs. S1C, S1D & S1E).
Cell death and DNA damage are more pronounced in AI glandsApoptotic cells with condensed nuclei started appearing in the lumens of the AI glands as early as day 8.5 PPM, while that was noted on and after day 17 PPM in the GI glands. TUNEL (terminal deoxynucleotidyl transferase dUTP nick end labeling) assay revealed a sharp increase in cell death within 36 h of pup withdrawal (AI-day 8.5 PPM) that peaked on day 11 PPM (5.1 ± 0.02% positive cells), followed by an equally sharp decline in cell death and return to near baseline by day 17 PPM (0.42 ± 0.0005% positive cells) (Fig. 2A and B). In contrast, there was a gradual increase in cell death in GI glands after day 12 PPM that peaked on day 25 (3 ± 0.1% positive cells), followed by a gradual decline to baseline by day 31 PPM (0.35 ± 0.0004% positive cells) (Fig. 2A and B). Overall, this shows that AI glands have significantly increased TUNEL-positive cells compared to the GI group averaged across all time points (p = 0.0005), predominantly due to the acute increase in cell death in AI glands between days 8.5 and 12.
While we observed a rapid increase in apoptosis in AI glands, the glands also demonstrated an increased activation of the DNA repair pathway. This pathway is one of the Hallmark gene sets [21] that we found differentially enriched in AI-derived LP cells harvested on day 28 PPM (negative enrichment score = 1.74, false discovery rate q-value = 0.0039, p-value = 0.024) when compared to GI-LP cells (Fig. 2C). This suggests an increased DNA damage response in AI glands. To investigate if indeed AI resulted in DNA damage that persisted over time, we analyzed the extent of DNA damage using γH2AX immunohistochemistry (IHC). There was a trend toward higher γH2AX-positive cells in AI vs. GI glands at day 120 PPM, although the difference was not statistically significant (Supplementary Fig. S2A, p = 0.10). Studies in C57BL/6 mice revealed a similar trend of increased DNA damage in AI-day 120 glands compared to GI-day 120 glands (p = 0.059, Fig. 2D and Supplementary Fig. S2B).
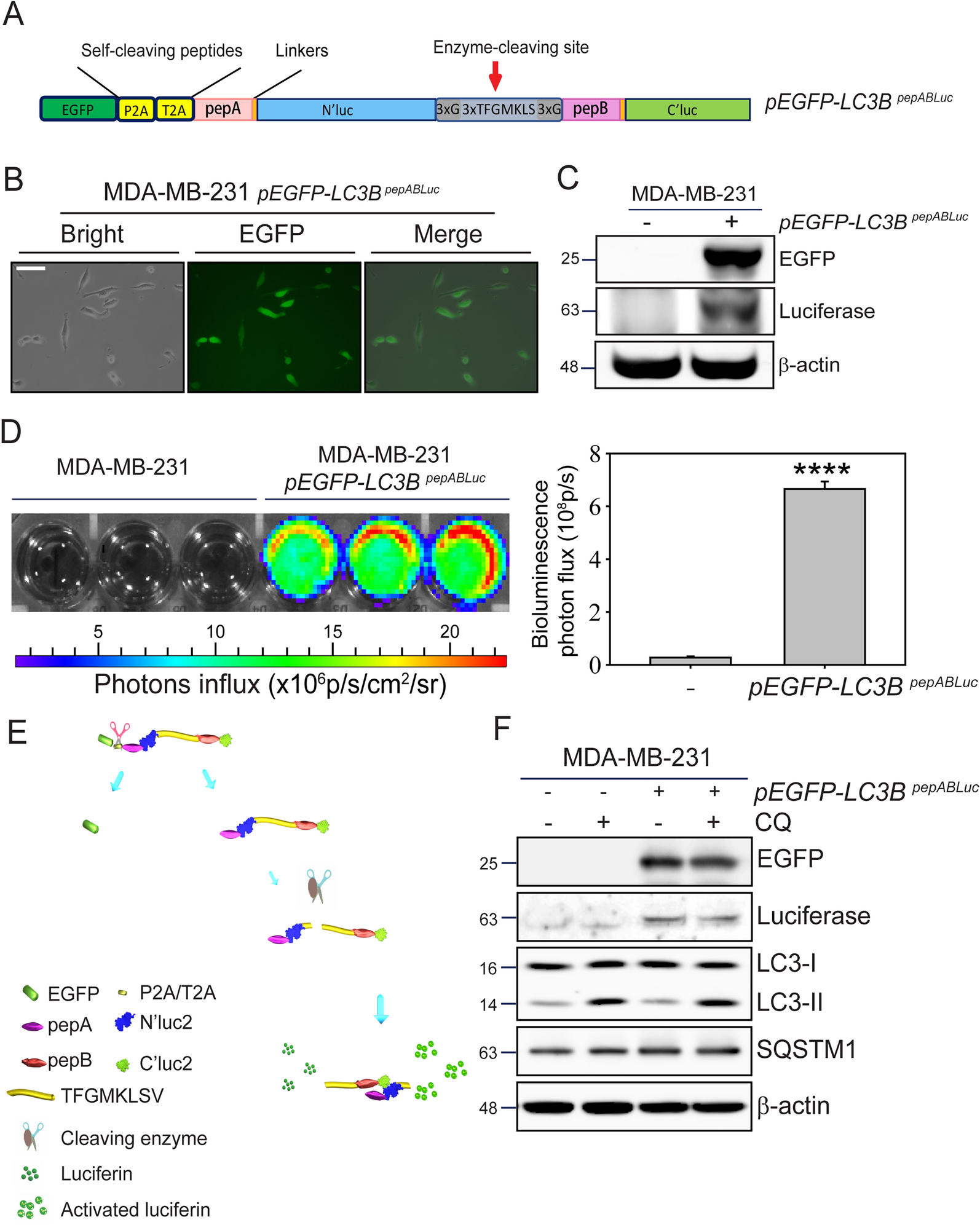
Fig.2
DNA damage response is heightened in AI glands compared to GI glands. A. Representative images of FFPE sections of AI and GI mammary glands subjected to TUNEL assay at indicated time points (day PPM). Arrows indicating apoptotic cells. B. Kinetics of cell death between days 7 and 28 PPM, assessed by TUNEL staining. n = 6 per group per time point. C. Enrichment plot of HALLMARK_DNA_REPAIR pathway in AI vs. GI glands on day 28 PPM. D. phospho-γH2AX positivity in, GI and AI glands harvested on day 120 PPM and in mammary glands of age-matched virgin mice. n = 4 per group. Ei. Western blot analysis of BCL-xL, BCL-xS and cleaved PARP in AI and GI mammary glands at early phase of involution, quantitated in Eii. BCL-xL, Eiii. BCL-xS and Eiv. 55 kDa cleaved PARP, and Ev. 24 kDa cleaved PARP
In involuting mammary glands, DNA damage initiates checkpoint mechanisms in mammalian cells, leading to cell cycle arrest to facilitate DNA damage repair. If the damage proves insurmountable, cell death is predominantly induced through apoptosis [22, 23]. Therefore, to study the apoptotic response in gradual and abruptly involuting mammary glands, we also analyzed BCL-xL, BCL-xS and the involvement of poly ADP-ribose polymerase (PARP). Levels of BCL-xL in AI glands are high at the onset of involution (day 8.5 PPM) and reduced to a low level (50% of day 7 PPM) on day 12. Although there was a slight increase in BCL-xL between days 17 and 25, surprisingly BCL-xL was very high on day 28 PPM (Figs. 2Ei & Eii). In contrast, we observed a relatively comparable level of BCL-xL at the early time points in GI glands, with the highest expression on day 25 followed by a marked reduction on day 28 (Figs. 2Ei and Eii). On the contrary, BCL-xS was detectable in the AI glands only on day 8.5 PPM. A consistent level of BCL-xS was present till day 22 PPM in the GI glands (Figs. 2Ei & Eiii). This demonstrates that in the AI glands, the key anti-apoptotic response was not as pronounced as in GI glands at the peak of cell death and that BCL-xS supporting mammary epithelial cell survival [24] was higher in both the glands until the onset of involution. Interestingly, the 89 kDa cleaved PARP, a product of caspase-mediated cleavage of PARP, was barely detectable either in the AI or GI glands of the FVB mice (Supplementary Fig. S2C). In contrast, a 55 kDa cleaved PARP, generated by cathepsin B-mediated cleavage [25], was detected in the GI glands; it gradually increased from day 17, peaked at day 25, and remained high on day 28 (Figs. 2Ei & Eiv). In AI glands, we observed a low level of 55 kDa cleaved PARP from day 11 PPM through day 28 PPM when compared to GI glands (Figs. 2Ei, Eiv & Supplementary Fig. S2D). A higher level of 24 kDa cleaved PARP, a trans-dominant inhibitor of PARP [26], was observed at the onset of AI (Figs. 2Ei, Ev & Supplementary Fig. S2D). Similarly, in the GI gland increased 24 kDa cleaved PARP level coincided with peak of cell death that remained high through day 28 PPM, suggesting a sustained inhibition of PARP. Collectively, our findings demonstrate a striking difference in the rate of cell death, adipocyte repopulation, PARP cleavage, and degree of DNA damage between the AI vs. GI glands.
Acute phase response genes are expressed primarily during AI but not during GIThe biphasic nature of postpartum mammary gland involution and identification of key players associated with acute phase response (APR) due to massive cell death stemmed from studies in mice subjected to AI following partum and brief lactation (7–10 days) [18, 27, 28]. However, the role of these genes during the gradual involution of mouse mammary glands remains unknown. Therefore, we next attempted to address whether there is a mere shift of key events and heightened expression of the APR genes during the peak of gradual involution (day 17–28 PPM) or alternatively if these changes are less pronounced, thus contributing to the protective effect against breast cancer risk. The fold change for all genes, both RNA and protein levels, at all the time points were measured relative to the levels on day 7 PPM (lactating gland).
In previous studies, STAT3 and C/EBPδ were identified as master regulators of lysosome-mediated program cell death (LM-PCD) [29, 30] using mouse models of AI. In our study, both Stat3 and C/ebpδ expression increased sharply and peaked on day 8.5 PPM (Stat3: 5.9-fold, p = 0.0007, C/ebpδ: 4.4-fold, p = 0.008) in the AI glands, followed by a rapid decline to near baseline by day 12 PPM. In contrast, GI glands showed a gradual increase in Stat3 expression that peaked on day 22 PPM (4.5-fold, p = 0.06) followed by a sharp decline on day 25 and thereafter (Figs. 3A and B). C/ebpδ expression was barely detectable in the GI glands throughout the process of involution. Western blot analysis demonstrated the highest level of Stat3 protein in the lactating glands that gradually declined with time in both AI and GI glands (Figs. 3Li & Liii). In contrast, the pStat3Y705 level in AI glands was highest (15-fold compared to day 7) and comparable at days 8.5 and 12 PPM, followed by a gradual decline until day 25. Interestingly, a second peak (~ sevenfold) was observed on day 28 PPM in AI glands. The pStat3Y705 level peaked in GI glands on day 22 PPM but was significantly less than that in AI glands at the onset of involution (sevenfold GI-day 22 vs. 15-fold AI-day 8.5; Figs. 3Li & Lii). Similarly, a sharp increase in pStat3Y705- positive cells on day 8.5 PPM (0.26% vs. 24.8% on days 7 and 8.5, respectively) was observed in the AI glands when mammary gland sections were analyzed by IHC (Supplementary Fig. 3A). This was followed by a 50% drop (11.9%) on day 12 PPM and further declined to 5.6% on day 22 PPM (Fig. 3M). Interestingly, an increase in pStat3Y705 positivity was observed on day 25 PPM (8.4%). In GI glands, pStat3Y705-positive cells were undetectable until day 12, followed by a gradual increase to 11.4% positivity on day 25 PPM (Fig. 3M). IHC analysis corroborates the data obtained by qPCR and Western blot analysis in both AI and GI glands.
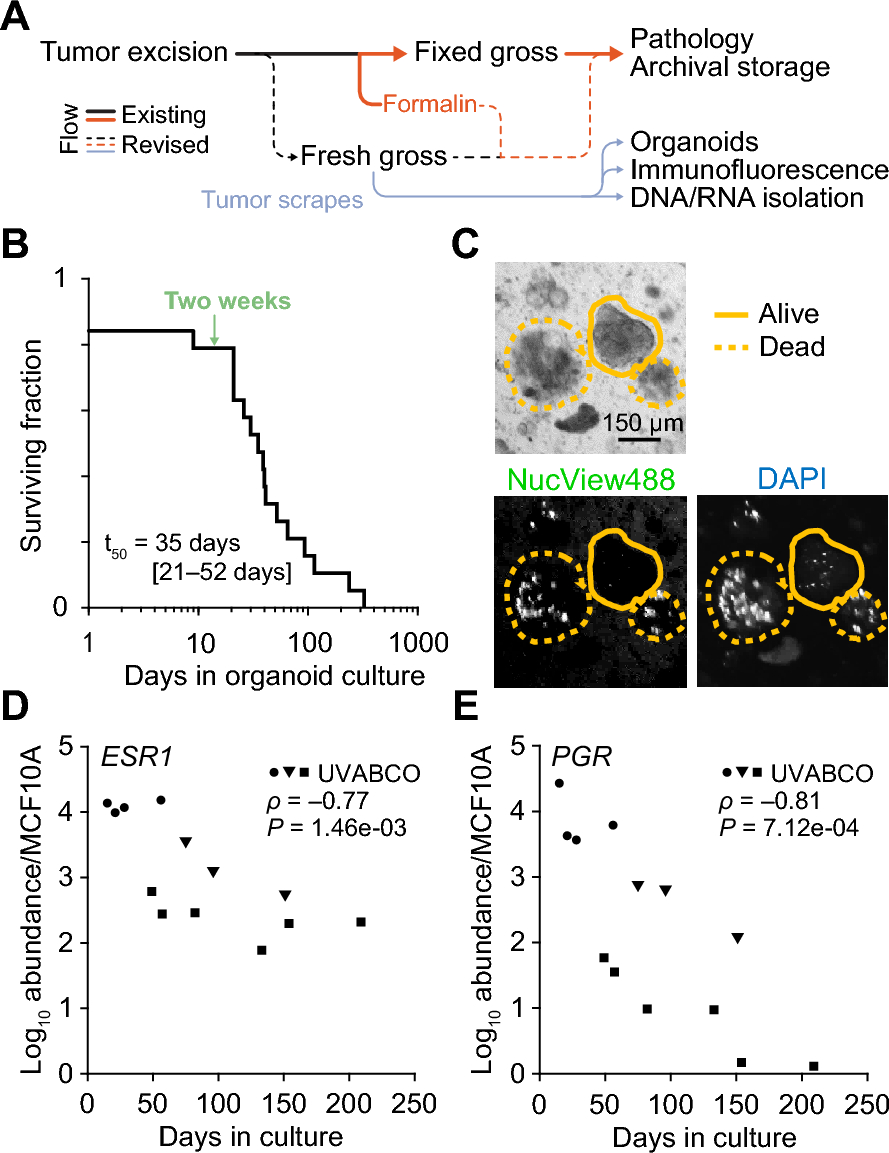
Fig.3
Expression pattern of acute phase response genes is distinct in AI and GI glands. Quantitative RT-PCR analysis of A. Stat3, B. C/EBPδ, C. CtsL, D. CtsB, E. Orm1, F. Orm2, G. Lrg1, H. Slpi, I. CD14, J. Chi3L1, and K. MMP2. Expression normalized to RPL37. n = 3 per group per time point. Li. Western blot analysis and quantitation of Lii. pStat3Y705, Liii. Stat3, Liv. CtsB, and Lv. CD14. M. Temporal pattern of pSTAT3Y705 positive cell distribution in AI and GI glands between day 7 and day 28 postpartum. n = 6 per group per time point. Ni. Representative images of FFPE sections of AI (day 11 PPM) and GI (day 22 and 25 PPM) mammary glands subjected to immunofluorescence analysis of pStat3Y705 (green) and CD14 (red). DAPI (blue) was used to counterstain the nuclei. Nii. Bar diagram showing the number of pStat3.Y705 and CD14 double-positive cells per field of view (FOV) in mammary gland sections of AI (day 11 PPM) and GI (day 22 and 25 PPM). n = 5 per group per time point. Number of FOV = 10
Analysis of CtsL and CtsB expression, the two lysosomal cysteine proteases involved in LM-PCD [27], revealed that in the AI glands, there was a sharp induction of CtsL (sixfold, p = 0.001) on day 8.5 PPM followed by a sharp decline on day 11 PPM. CtsL expression remained low at the subsequent time points (Fig. 3C). There was no marked change in CtsL expression in the GI glands, suggesting a minimal role in cell death during GI (Fig. 3C). Intriguingly, in the AI glands CtsB expression showed biphasic expression with the first peak at day 12 PPM (3.5-fold., p = 0.01), followed by a gradual decline through day 17 and an increase again on day 22 (2.5-fold., p = 0.05) (Fig. 3D). GI glands did not exhibit this biphasic expression and rather showed a gradual increase over time, which peaked on day 25 (2.6-fold, p = 0.04) (Fig. 3D). The increased level of CtsB expression was concurrent with maximum cell death in the GI glands, suggesting a major role of CtsB and not CtsL in cell death during GI. We also observed the highest level of CTSB protein (27 kDa) in AI-day 11 glands (~ sixfold higher compared to day 7), which started to increase on day 8.5 and decreased after day 12. CTSB protein peaked on day 22 in GI glands (a twofold increase over day 7) and remained elevated through day 28 (Figs. 3Li & Liv, Supplementary Fig. S3B).
Several downstream effectors of Stat3 and predominantly APR/immunomodulatory proteins have been identified to play a key role during mouse mammary gland involution in the AI setting [18]. We analyzed the expression of these key players (Orm1, Orm2, Lrg1, and Slpi) to assess their role in GI. Both Orm1 and Orm2, the two APR genes, showed a sharp increase in expression in the AI glands following the removal of pups, with the highest expression on day 12 PPM (Orm1: tenfold, p = 0.04; Orm2: 370-fold, p = 0.05) and a sharp decline to baseline by day 17 PPM (Fig. 3E and F). In GI glands, Orm1 expression was highest at day 28 (2.2-fold, p = 0.5), while Orm2 was not detectable (Fig. 3E and F). Similarly, expression of Lrg1, a late-stage macrophage marker, showed a marked increase on day 12 PPM in AI glands (203-fold, p = 0.01), followed by a sharp decline to baseline by day 17 PPM and remained low (Fig. 3G). In GI glands, expression of Lrg1 was undetectable throughout with a small increase on day 25 (28-fold, p = 0.02) (Fig. 3G). Slpi, a secretory leukocyte peptidase inhibitor is an immunomodulatory protein that protects tissue epithelium from serine proteinases [31]. AI glands demonstrated a sharp increase in Slpi expression that peaked on day 8.5 PPM (61-fold, p = 0.008), followed by a decline to baseline by day 12 PPM, whereas its expression was undetectable in GI glands throughout the involution process (Fig. 3H). In brief, these results suggests that while an acute phase response is obvious during AI, GI glands do not display such a response, even at the peak of cell death (day 25 PPM, Fig. 2B).
A key finding by Stein et al. was the significantly high expression of CD14 by luminal epithelial cells and their potential role in dead cell clearance immediately after the removal of pups in the AI setting [19]. Anticipating a similar role of luminal epithelial cells in GI glands to coincide with maximum cell death, we compared CD14 expression at both RNA and protein levels between GI and AI glands over time. Our data shows a 20-fold increase in CD14 expression in the AI glands from day 7 to day 8.5 PPM (p = 0.005), and a sharp decline by day 11 PPM (Fig. 3I). In contrast, only a very low level of CD14 expression was observed in the GI glands on day 25 PPM (3.4-fold, p = 0.003, Figs. 3I). Western blot analysis shows that CD14 protein levels followed a similar pattern as gene expression in both GI and AI glands (Figs. 3Li & Lv, Supplementary Fig. S3C). In the AI glands, the CD14 protein level was highest on day 8.5 PPM (25-fold increase), and in the GI glands, CD14 protein level was highest on day 22 (sixfold increase). We also performed immunofluorescence analysis on AI glands at day 11 PPM and GI glands at days 22 as well as day 25 PPM to assess the presence of alveolar clusters co-expressing phosphorylated Stat3 (pStat3Y705) and CD14. Our analysis revealed a significantly higher number of pStat3 + CD14 + cells in the AI glands at day 11 PPM compared to GI glands at days 22 or 25 PPM (Figs. 3Ni & 3Nii, p<0.001). Therefore, this data suggests a minimal role of luminal epithelial cells in the clearance of apoptotic cells during GI as opposed to AI.
We next analyzed Chi3L1 expression, a tissue-restricted marker of late-stage macrophage differentiation involved in wound healing. Increased expression of this gene was reported by Scully et al. during mammary gland involution in the AI setting [32]. Our data shows that Chi3L1 expression in AI glands peaked on day 11 PPM (555-fold, p = 0.002) and remained high until day 22, which was followed thereafter by a gradual decline until day 28 (Fig. 3J). In the GI glands, Chi3L1 gradually increased after day 22 PPM and peaked on day 25 (750-fold, p = 0.03). Expression of MMP2, a matrix metalloprotease involved in the restructuring of post-involution glands [33], was high in AI glands as early as day 11 PPM and remained high until day 22 PPM, while it was upregulated in GI glands around day 25 PPM (Fig. 3K).
Altogether, based on gene and protein expression analysis of the whole mammary glands, we see diverse and distinct patterns between AI and GI glands, which are summarized in Table 1.
Table 1 Expression pattern of acute phase response genesMyeloid-derived suppressor cells and M2-like macrophages release CCL9 in AI glands at the peak of cell deathOur previous study demonstrated that inflammation persists in the mammary glands that had undergone AI, whereas it is transient in the GI glands with a quick return to baseline [13]. Here we examined the kinetics of immune cell infiltration at the early stages of AI vs. GI, specifically, at the peak of cell death. Analysis of macrophage infiltration by IHC (F4/80) revealed a sharp increase in AI glands following removal of the pups that peaked on day 11 PPM (35 ± 8%) and gradually declined to ~ 14% positivity on day 17 PPM. Interestingly, there was a second surge of macrophages (~ 23% positive cells) on days 22 and 25 PPM (Fig. 4A and Supplementary Fig. S4). In contrast, macrophage infiltration in GI glands remained low until day 12 PPM, followed by a slow and gradual increase over time to 27 ± 7% positivity on day 25 PPM. Statistical analysis shows that AI has significantly increased F4/80 staining compared to the GI group averaged across all the time points (p = 0.0005) by 6.62%; this was mainly attributed to the differences at the earlier time point (days 11 and 12 PPM). This data demonstrates a biphasic pattern of macrophage infiltration in the AI gland, an acute inflammatory response at the onset of involution, and a second surge later around day 25 PPM. Intriguingly, our previous study demonstrated a decline in F4/80 positive cells in GI glands at later time point (day 56 PPM), while it continued to remain high in the AI glands [13]. Thus, in the GI gland, the macrophage infiltration is more restricted coinciding with cell death and declines once involution is complete while in the AI glands it is sustained beyond cell death.
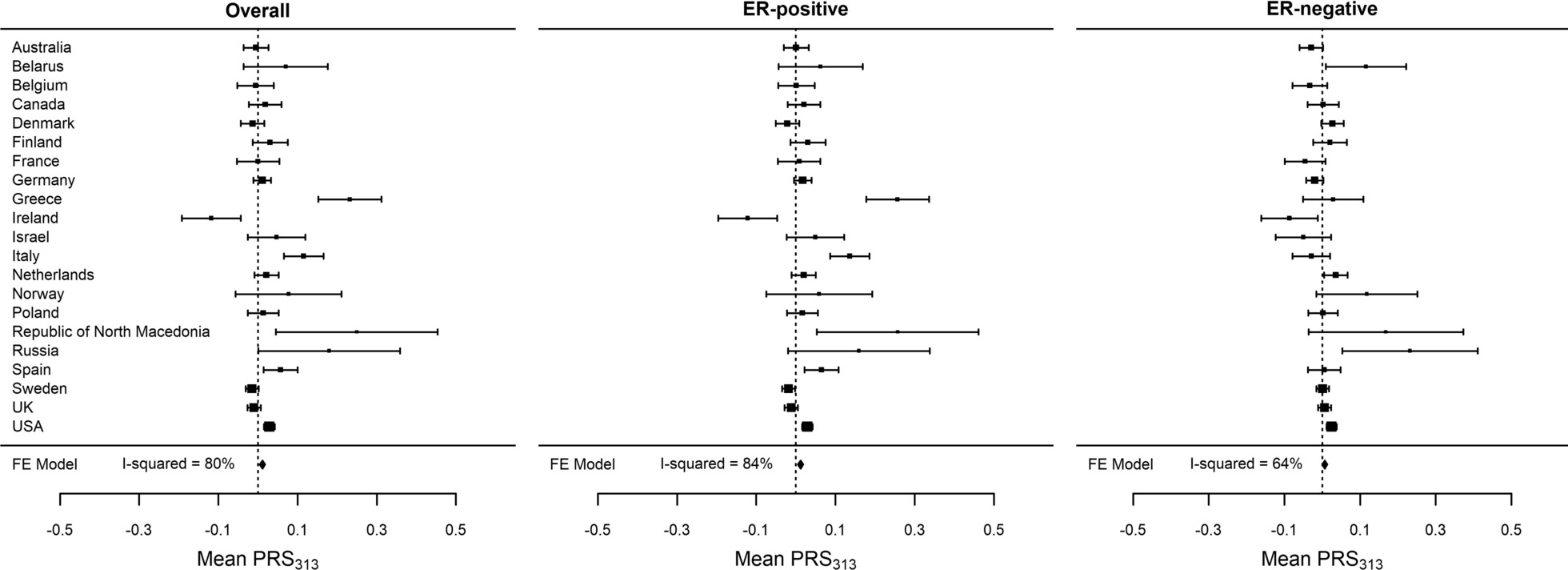
Fig. 4
Abundance of myeloid-derived suppressor cells and M2-like macrophages in AI glands at the peak of cell death secreting CCL9. A. Temporal pattern of F4/80 positive cell distribution in AI and GI mammary glands harvested between days 7 and 25 PPM. FFPE sections of mammary glands were subjected to IHC using F4/80 antibody. Images of the stained slides were captured using Vectra 3.0 and analyzed using InForm software. n = 6 per group per time point. B. Mammary glands from AI-day 11 and GI-day 25 female mice were processed to identify different subpopulations of myeloid cells by multi-color flow cytometry. The representative flow cytometric dot plots and graphs of % B. CD45 + leukocytes; C. CD11b + Gr1 + MDSCs (out of CD45 + cells); D. CD11b + F4/80 + total macrophages (out of CD45 + cells); E. MHCII + M1-like macrophages (out of CD45 + CD11b + F4/80 + total macrophages); F. CD206 + M2-like macrophages (out of CD45 + CD11b + F4/80 + total macrophages); n = 3 per group. G. CD11b + Gr1 + MDSCs, and H. CD11b + F4/80 + MHCII-CD206 + M2-like macrophages were sorted from mammary glands of AI-day 11 and GI-day 25 mice. The cells were lysed and subjected to cytokine/chemokine array analysis. The images show expression of cytokine/chemokine and bar diagrams showing CCL9 expression quantified using ImageJ software. I. Quantitative RT-PCR analysis of Ccl9 expression in mammary glands over time (days 7 to 28 PPM), n = 3 per group. J. Quantitative RT-PCR analysis of Tgfβ3expression in mammary glands over time (days 7 to 28 PPM), n = 3 per group
Next, we assessed the myeloid cell composition during the peak of cell death in both gland types based on previous reports showing the recruitment of various myeloid cell subsets in the developing and inflamed mammary gland [34]. Their relative abundance varies during post-lactational regression and contributes to a microenvironment conducive to tumor growth [35]. However, a comparative analysis of myeloid cell distribution in AI and GI mammary glands has not been conducted so far. Histological examination of H&E-stained sections revealed a significant increase in inflammatory cell infiltration starting on day 11 in AI glands and day 25 in GI glands, coinciding with the respective cell death peaks (Figs. 1 & 2).
To further analyze myeloid cell populations, we performed multi-color flow cytometric analysis on AI-day 11 and GI-day 25 mammary glands. The relative abundances of total CD45 + leukocytes and CD11b + F4/80 + total macrophages were not statistically different between these two groups (Fig. 4B and D). However, we observed a significant enrichment of CD11b + Gr1 + myeloid - derived suppressor cells (MDSCs) in mammary glands from AI-day 11 compared to GI-day 25 (Fig. 4C), while MHCII + M1-like macrophages did not exhibit a significant change between the two groups (Fig. 4E). On the other hand, CD11b + F4/80 + CD206 + cells that were identified as M2-like macrophages (pro-tumor) [36] were more abundant in AI-day 11 compared to GI-day 25 glands (Fig. 4F). To further characterize the functional properties of macrophage subpopulations, Gr1 + MDSCs and CD206 + M2-like macrophages were sorted from AI-day 11 and GI-day 25 glands and subjected to mouse cytokine/chemokine array analysis. Both MDSCs and M2-like macrophages from AI glands showed a significant increase in the expression of CCL9 compared to their counterparts from GI-day 25 mammary glands (Fig. 4G and H). Additionally, qRT-PCR validation demonstrated significantly higher Ccl9 expression in AI glands, however, showing a biphasic expression (Fig. 4I). Previous studies have shown that TGFβ3 promotes the recruitment of inflammatory cells by upregulating the production of chemokines/cytokines such as CCL9 [37]. Additional studies reported TGFβ3 upregulation and its active involvement during mammary gland involution [38, 39]. To connect the dots, we measured the expression of Tgfβ1, Tgfβ2, and Tgfβ3 in GI and AI glands. Importantly, our data shows significantly increased expression of only Tgfβ3 that coincides with cell death peak in both AI and GI glands (Fig. 4J). In the AI glands, Tgfβ3 expression is elevated again around day 22 PPM. The marked overlap in the biphasic pattern of Ccl9 and Tgfβ3 expression in the AI glands suggests a possible role of TGFβ3 in regulating Ccl9 expression. Conversely, Tgfβ1 and Tgfβ2 expression was undetectable in both AI and GI glands. Taken together, these results show that at the peak of cell death, AI glands have more MDSCs and M2-like macrophages compared to GI glands, concomitant with elevated levels of CCL9 chemokine.
CCL9 promotes the expansion of ERα negative luminal progenitor cellsWe previously reported the expansion of LP cell populations in AI but not GI mammary glands [13]. To further investigate the impact of CCL9 on LP cells, we cultured sorted LP cells from 6-week-old virgin female mice as 3D-organoids and treated them with mouse recombinant CCL9 (100 ng/mL). Three-dimensional organoids formed by different cell types recapitulate specific morphologies in vitro [40], and both mammary luminal and basal cells are maintained faithfully in 3D assays when cultured for a short time [41]. LP cells cultured on basal medium eagle (BME) without CCL9 treatment formed solid round organoids. In contrast, on day 12 of CCL9 treatment, we observed the formation of disorganized acinar-type organoids (Fig. 5A, panel 3), with a 6.6-fold increase in luminal budding and branching outgrowths (Fig. 5B, p < 0.0001). Interestingly, similar morphological differences were observed in LP cells isolated from AI and GI mammary glands on day 56 PPM and grown as organoids (Fig. 5C). A recent study demonstrated that basal/stem cells can form these disorganized organoids with luminal characteristics [40], suggesting that LP cells cultured in vitro may acquire basal phenotypes. To understand the effect of CCL9 on LP cells, we analyzed the expression of several genes associated with stemness (CD133, CD49f, CD44), proliferation (Ki67), basal vs. luminal characteristics (Krt14, Krt8), LP cell markers (Elf5) [42], and Esr1. Control organoids (untreated) and the organoids treated with CCL9 for 12 days as described previously (Fig. 5A) were used for RNA isolation and subjected to qRT-PCR. CCL9 treatment of the LP organoids for 12 days resulted in a significant increase in Elf5 expression (twofold, p = 0.004), and a marked reduction in Esr1 expression (−2.5-fold, p = 0.001) (Fig. 5D and E). Although there was a trend toward increased Ki67 expression after 12 days of CCL9 treatment (Supplementary Fig. S5A), no significant changes were observed in the expressio
留言 (0)