
記住我


To study the potential role of KDM6A at the onset of XCI, we used CRISPR/Cas9 to knockout (KO) Kdm6a in hybrid female mouse ES cells derived from a 129 x Mus castaneus (cast) cross in which alleles can be distinguished by SNPs (Additional File 1: Figure S1A). In order to examine changes specifically on the Xi we used Tsix-stop cells (donated by J. Gribnau, Erasmus MC) in which a transcriptional stop signal is inserted onto the 129 allele of Tsix, resulting in completely skewed silencing of the 129 X chromosome upon differentiation [18]. Stable Kdm6a KO Tsix-stop ES cell clones with a heterozygous or homozygous deletion of part of exon 2 through part of exon 4 (wt/ΔE or ΔE/ΔE, respectively) were generated (Additional File 1: Figure S1A) [14]. PCR and Sanger sequencing verified homozygous editing of Kdm6a in two KO clones (Tsix-Kdm6aΔE/ΔE17 and Tsix-Kdm6aΔE/ΔE21) and heterozygous editing in one KO clone (Tsix-Kdm6awt/ΔE4). Despite residual Kdm6a expression in the homozygous KO clones, western blot analysis showed no evidence of KDM6A protein [14, 15]. We also isolated control Tsix-stop ES cell clones (Tsix-cln1, Tsix-cln2; hereafter called CRISPR controls), which were subject to the CRISPR/Cas9 treatment but did not exhibit a deletion. In a separate experiment, Kdm6a homozygous editing was done by deleting the promoter in another hybrid female mouse ES cell line called E8 (donated by J. Gribnau, Erasmus MC) to derive clone E8-Kdm6aΔP/ΔP13 (Additional File 1: Figure S1A). The E8 cell line derived from a C57BL/6J (BL6) x cast cross undergoes random XCI with partial skewing due to a strong Xce allele that results in a ~2:1 inactivation of the BL6:cast X chromosome upon differentiation [4, 19]. A summary of the clones used in this study can be found in Additional File 2: Table S1.
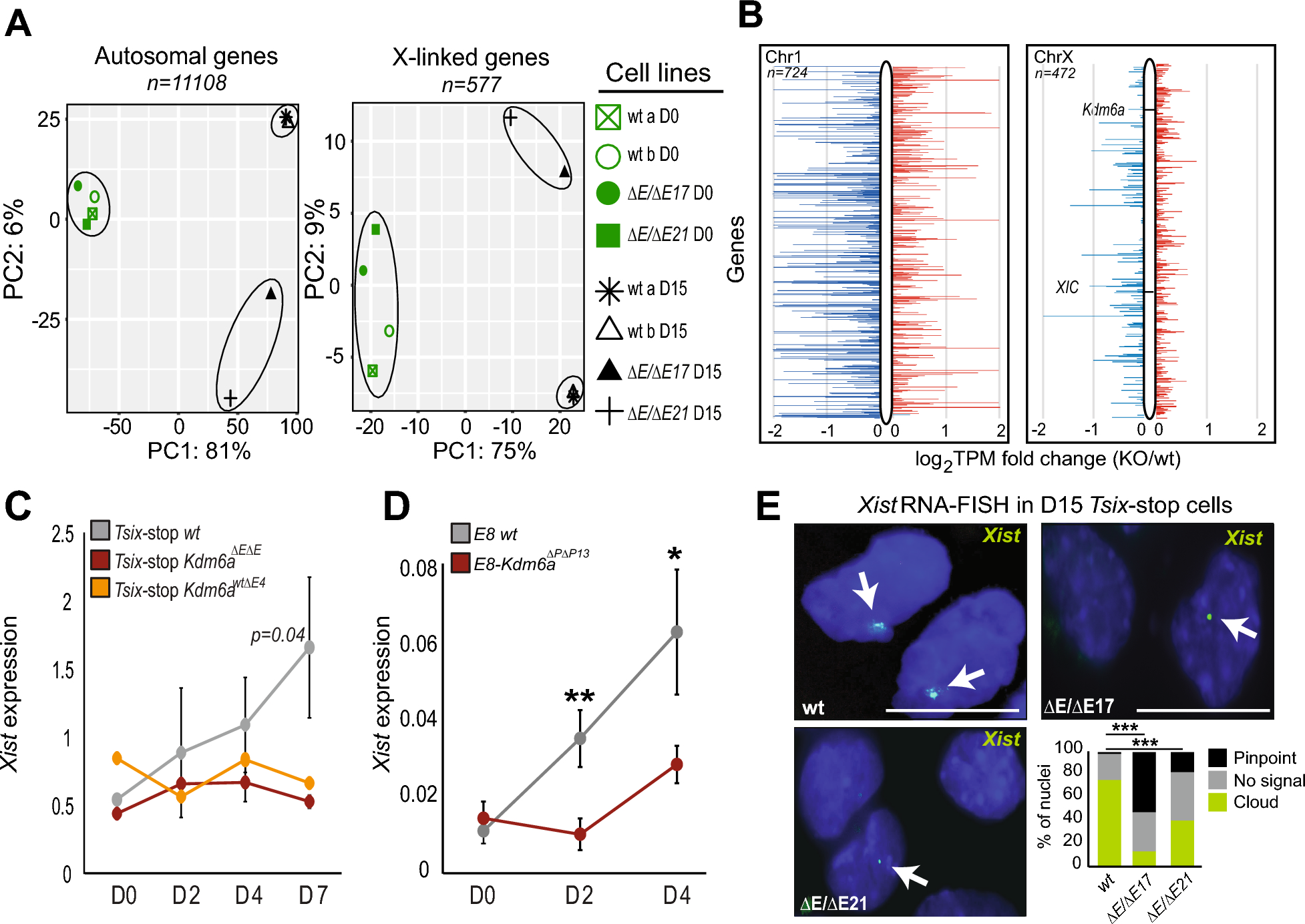
Kdm6a KO results in expression changes of genes involved in germ layer development after XCI occursRNA-seq analyses were done to compare gene expression between Tsix-stop wild-type (wt) and Kdm6a KO (Tsix-Kdm6aΔE/ΔE17 and Tsix-Kdm6aΔE/ΔE21) clones before day 0 (thereafter abbreviated D0) and after (D15) differentiation of ES cells into embryoid bodies (EB). For Tsix-stop cells, principal component analysis (PCA) based on either autosomal or X-linked gene expression shows little separation between wt and KO clones at D0, but a clear separation at D15, indicating that the effects of Kdm6a KO are most pronounced after differentiation (Figure 1A). Consistent with PCA clustering, we found only 624 differentially expressed genes (DEGs) between wt and KO cells at D0, while there were 886 DEGs at D15 (≥2 fold change; FDR ≤0.1) (Additional File 1: Figure S1B; Additional File 3: Table S2). These findings are in agreement with KDM6A’s dispensable role in undifferentiated ES cells versus its multiple roles in regulation of genes involved in development and differentiation [11, 14, 20]. Indeed, among the DEGs found at D15 (a time point after XCI has occurred) there is a subset of genes involved in germ layer development, including genes expressed in all three germ layers, which are downregulated in Tsix-stop Kdm6a KO cells at D15, but not at D0 (Additional File 1: Figure S1C; Additional File 4: Table S3). Of the germ layer genes with decreased expression in Tsix-stop KO cells, 19 were expressed (>1 TPM) at D2 and 18 were expressed at D4 of differentiation in E8-Kdm6a wt cells. There was no clear pattern of downregulation of these genes at either time point following Kdm6a KO, strongly suggesting that germ layer gene downregulation occurred after XCI (Additional File 1: Figure S1C; Additional File 4: Table S3). In line with KDM6A’s role in facilitating gene expression by removing the repressive mark H3K27me3, we observed an overall decrease in autosomal gene expression (Figure 1B). In contrast, 217/472 (46%) of X-linked genes assayed showed increased expression (>1.25 TPM fold change) in Tsix-stop Kdm6a KO (average of clones Tsix-Kdm6aΔE/ΔE17 and Tsix-Kdm6aΔE/ΔE21) compared to wt at D15 (Figure 1B).
Fig. 1
Kdm6a KO results in Xist downregulation. A Principal component analysis (PCA) based on expression of autosomal or X-linked genes in two Tsix-stop wt clones and two Kdm6a KO clones (Tsix-Kdm6aΔEΔE17 and Tsix-Kdm6aΔEΔE21) before (D0) and after differentiation (D15). Only genes with a minimum of 2.5 CPM in 4 libraries were considered. PCA shows separation of clones based on Kdm6a KO status mainly after differentiation. PCA figures were generated by iDEP.95. B Plots of diploid gene expression from a representative autosome (Chr1, left) with similar gene content as the X, and from the X chromosome (right) in Tsix-stop wt and KO clones (average for Tsix-Kdm6aΔEΔE17 and Tsix-Kdm6aΔEΔE21) at D15. Kdm6a and the XIC are highlighted. Log2 TPM fold change (KO/wt) is shown. Genes with a decrease of expression in KO versus wt are in blue, and with an increase in red. C qRT-PCR of average Xist expression normalized to Actinβ during ES differentiation in three Tsix-stop wt samples, two Kdm6aΔEΔE KO clones, and one Kdm6aΔwtΔE clone. p-value is derived from comparing wt to Kdm6aΔEΔE biological replicates. Divergence of Xist expression between wt and KO cells begins at D2 and is largest at D7 (p = 0.04). The Kdm6awt/ΔE4 heterozygous KO clone behaves similarly to the homozygous clones. D qRT-PCR of Xist expression in E8 wt and KO clone (E8-Kdm6aΔPΔP13). Expression is normalized to Actinβ. Xist is significantly lower at D2 and D4 in KO cells. P-values derived from two technical replicates of wt and KO cells (**p < 0.01;*p < 0.05). E RNA-FISH with a probe specific for Xist RNA labeled in green in Tsix-stop wt and KO clones (Tsix-Kdm6aΔEΔE17 and Tsix-Kdm6aΔEΔE21). Examples of nuclei are shown together with histograms for quantification of signal type (pinpoint, no signal or cloud). Xist clouds are present in 74% of D15 wt cells, but significantly decrease in KO clones (cloud compared to no signal; ***p < 0.00001 for Tsix-Kdm6aΔEΔE17 and < 0.0001 for Tsix-Kdm6aΔEΔE21 fisher’s exact test), while the frequency of pinpoint Xist signals increases in KO clones (pinpoint in wt compared to pinpoint in KO; ***p < 0.00001 for Tsix-Kdm6aΔEΔE17 and Tsix-Kdm6aΔEΔE21 fisher’s exact test). Nuclei are counterstained with Hoechst 33,342. Scale bar = 10 µm
Xist expression and coating are impaired during early differentiation following Kdm6a KOOne of the earliest events at the onset of XCI is the upregulation of Xist [1, 2]. Therefore, we investigated changes in the dynamics of Xist expression during early mouse ES cell differentiation at D0, D2, D4, D7, and D15 following Kdm6a KO. Compared to Tsix-stop wt and CRISPR-control clones, Kdm6a KO clones (Tsix-Kdm6aΔE/ΔE17 and Tsix-Kdm6aΔE/ΔE21) have significantly reduced levels of Xist expression beginning at D2 of differentiation when Xist upregulation normally starts (Figures 1C and Additional File 5: Figure S2A). Importantly, this finding was confirmed in Kdm6a KO clone (E8-Kdm6aΔP/ΔP13) derived from an independent ES line E8 (Figure 1D). Thus, we infer that KDM6A contributes to the activation of Xist in early differentiation beginning prior to D2 and increasing at D4 and D7 (Figures 1B and Additional File 5: Figure S2B). Analysis in Tsix-stop cells with heterozygous deletion of Kdm6a (Tsix-Kdm6a wt/ΔE4) revealed a similar effect on Xist by D7, suggesting that expression of Kdm6a from both alleles is needed for activation of Xist (Figure 1C). The lower expression of Xist in Kdm6a KO cells does not appear to be due to changes in levels of several of the known Xist repressors (Tsix, Nanog, Pou5f1, Sox2, Rex1) or activators (Yy1, Rlim) as these were not differentially expressed between Tsix-stop wt and KO cells at D0 (Additional File 5: Figure S2C) [21]. Klf4, which is known to repress Xist via activation of Tsix [22], did show an increase in Kdm6a KO cells at D0, but Tsix expression was consistently lower in KO cells at D2, reaching levels similar to those in wt only at D4, suggesting that the increase in Klf4 did not affect Tsix expression at this stage (Additional File 5: Figure S2D and E; Additional File 3: Table S2). Note that despite a small increase in Sox2 expression in Kdm6a KO cells, this gene was not called as a DEG by Deseq2 (Additional File 5: Figure S2C; Additional File 3: Table S2). Previous studies have shown aberrantly increased expression of Xist with Tsix inhibition in undifferentiated ES cells grown in media with serum [23, 24]. However, we verified that the Tsix-stop ES cells used here maintain a low level of Xist expression at D0 (~2 TPM) similar to levels observed in previous studies despite the presence of serum in the media (Additional File 5: Figure S2F) [3, 14].
To determine whether Kdm6a KO affects the formation of an Xist cloud we performed RNA-FISH with a probe that targets Xist in differentiated Tsix-stop Kdm6a KO and wt cells at D15. The percentage of nuclei with Xist clouds sharply decreased after KO, with only 18% and 38% of nuclei with an Xist cloud in clones Tsix-Kdm6aΔE/ΔE17 and Tsix-Kdm6aΔE/ΔE21, respectively, versus 74% of wt nuclei (Figure 1E). Consistent with reduced Xist expression in Kdm6a KO clones, nuclei with a small pinpoint Xist signal rather than a cloud were prevalent in these clones (41% and 23% of nuclei in clones Tsix-Kdm6aΔE/ΔE17 and Tsix-Kdm6aΔE/ΔE21, respectively) compared to 4% in wt nuclei (Figure 1E). Loss of an X chromosome during cell culture is a common occurrence in female mouse ES cells and would confound our analysis of X-linked gene expression. To address this possibility DNA-FISH using probes targeting X chromosome-specific regions was done to verify that minor X chromosome loss occurred in any condition tested (Additional File 6: Figure S3).
Together, these results indicate that KDM6A contributes to upregulation of Xist expression in a threshold dependent manner, and to Xist coating of the Xi during early mouse ES cell differentiation.
Aberrantly increased gene expression from the Xi in differentiated Kdm6a KO cellsWe postulated that reduced Xist upregulation and Xist coating in Kdm6a KO cells may lead to impaired XCI in these cells. Allelic analyses of X-linked gene expression in Tsix-stop cells show that the increase in expression following Kdm6a KO is mainly due to increased expression specifically from the Xi where 76% (242/317) of assayed genes show a ≥ 1.25 TPM fold increase (Figure 2A, B; Additional File 8: Table S4). In contrast, there is no clear directionality for dysregulated genes on the Xa (54% up and 46% downregulated; ≥ 1.25 TPM fold cutoff) (Figure 2C). In addition, we confirmed increased X-linked gene expression in an independent Kdm6a KO clone (E8-Kdm6aΔP/ΔP13) where 42% (208/491) of X-linked genes show a ≥ 1.25 TPM fold increased expression at D4 of differentiation (Additional File 7: Figure S4; Additional File 8: Table S4). In E8 cells, where XCI is largely random, allelic analyses show increased X-linked gene expression following Kdm6a KO coming from the BL6 and cast X (Additional File 7: Figure S4A, B). However, gene expression is slightly more elevated from the BL6 X, suggestive of a specific effect of Kdm6a KO on the Xi, which would tend to be the BL6 X due to partial XCI skewing in E8 cells (Additional File 7: Figure S4C, D). Thus, failure to upregulate Xist leads to increase X-linked gene expression in differentiated cells. Considering autosomal genes, Kdm6a KO resulted in a majority of dysregulated genes with decreased expression (84% and 60% of dysregulated genes in Tsix-stop Kdm6a KO clones and in E8-Kdm6aΔP/ΔP13, respectively), consistent with the role of KDM6A in facilitating gene expression through removal of H3K27me3 (Additional File 3: Table S2).
Fig. 2
Kdm6a KO leads to increased gene expression from the Xi in differentiated cells. A Allelic X-linked gene expression ratios (129:cast) in Tsix-stop wt, CRISPR control, and KO cells (Tsix-Kdm6aΔEΔE17 and Tsix-Kdm6aΔEΔE21) at D0 and D15 of differentiation. Ratios between expressed X-linked genes (> 1TPM) are shown. There is increased gene expression from the 129 X-chromosome (Xi) following Kdm6a KO differentiation. B, C Plots of expression from the Xi (B) and the Xa (C) (average for Tsix-Kdm6aΔEΔE17 and Tsix-Kdm6aΔEΔE21) at D15. Kdm6a and the XIC are highlighted. Log2 TPM fold change (KO/wt) is shown. Genes with a decrease of expression in KO versus wt are in blue, and with an increase in red. Gene expression from the Xi is selectively increased following KO. D Top, scatter plots of average log2 expression between Tsix-stop wt and KO clones (average for Tsix-Kdm6aΔEΔE17 and Tsix-Kdm6aΔEΔE21) at D15 for X-linked genes categorized as those silenced early, silenced at an intermediate time, silenced late, and not silenced (escape) during XCI [3, 25]. KO clones show higher expression of genes in each category compared to wt. n indicates the number of genes in each category. Bottom, histograms of median allelic expression of genes silenced at different times during XCI and of genes that escape XCI is increased in Tsix-stop KO clones compared to wt and CRISPR controls for all categories, while there is little change in expression from the Xa. n indicates the number of genes in each category. E Expression fold changes of individual escape genes from the Xi in Tsix-stop KO clones (average for Tsix-Kdm6aΔEΔE17 and Tsix-Kdm6aΔEΔE21) compared to wt at D15. Most genes show increased expression
Next, we grouped X-linked genes in four defined temporal clusters, early, intermediate, late, and not silenced (or escape), based on their previously reported timing of silencing during differentiation [3, 25]. All gene clusters show a trend of increased expression from the Xi in Tsix-stop Kdm6a KO cells compared to wt or CRISPR controls (Figures 2D, E; Additional File 9: Table S5). Interestingly, the expression of most escape genes is also increased after Kdm6a KO (Figure 2D, E). For all clusters the increase is usually less than ~ 1.25 fold, with few escape genes showing expression changes > 1.25 fold in KO versus wt ES cells. Few escape genes show a decrease in expression and include Xist and Kdm6a, as expected (Figure 2E).
Taken together, these results indicate a reduced potency of XCI in differentiated Kdm6a KO cells, which implicates KDM6A, at least in part, as a regulator of X-linked gene expression dosage via upregulation of Xist during the onset of XCI.
Kdm6a KO results in dysregulated H3K27me3 levels on the X chromosomeNext, we investigated changes in H3K27me3 profiles using Cut&Run to compare wt Tsix-stop cells to Kdm6a KO clone Tsix-Kdm6aΔE/ΔE17. At D0 and D15, H3K27me3 levels (H3K27me3 peak numbers) appear similar along the whole X chromosome between wt and Kdm6a KO (Figure 3A), but a closer inspection revealed changes at the Xist locus. Initially (D0) H3K27me3 enrichment is only slightly higher across the Xist gene in Kdm6a KO ES cells, consistent with KDM6A’s role in resolving bivalent chromatin (Figure 3B). However, following XCI at D15, there is a marked increase in H3K27me3, which correlates with lower Xist expression that culminates at ~ 2.5 fold upregulation between D0 and D15 in KO cells, compared to ~ 6.0 fold in wt cells (Figure 3B, C). In addition to changes at Xist, H3K27me3 enrichment is also markedly higher at Cdx4, the Tsix/Xist locus, and the Ftx 5’ region in Kdm6a KO versus wt cells at D15 (Additional File 10: Figure S5A). Both Cdx4 and Ftx show little expression change following Kdm6a KO, however, these genes are expressed at very low levels following differentiation (< 1TPM) and thus are not classified as DEGs (Additional File 10: Figures S5B–E). As a control, we analyzed a subset of known KDM6A target genes where we confirmed an increase in H3K27me3 levels associated with decreased expression in KO versus wt cells (Additional File 11: Figure S6A). Genes without significant expression change in KO cells did not show differences in H3K27me3, as expected (Additional File 11: Figure S6B). Consistent with our previous study, the imprinted Dlk1/Meg3 locus showed an increase H3K27me3 levels in Kdm6a KO cells, which is specifically located along the imprinting control regions (ICR), and concordant with decreased gene expression (Additional File 11: Figure S6C; Additional File 3: Table S2) [14].
Fig. 3
KDM6A binds to the promoter of Xist and Kdm6a KO results in increased H3K27me3. A IGV browser view of H3K27me3 peak profiles across the entire X chromosome in wt Tsix-stop cells (blue) and KO cells (Tsix-Kdm6aΔEΔE17) (red) at D0 and D15. The number of X chromosome peaks for each condition and time point are in parentheses. Green bars underneath represent genes along the X chromosome. The Xist gene is marked. B IGV browser view of H3K72me3 profiles and peaks at the Xist locus in wt Tsix-stop (blue) and KO Tsix-Kdm6aΔEΔE17 cells (red) at D15. There is a slight increase in H3K27me3 levels in KO cells at D0, and a marked increase at D15. C Histogram of log2 fold change of Xist expression in wt (2 isolates: wt A, wt B) and two KO clones (Tsix-Kdm6aΔEΔE17 and Tsix-Kdm6aΔEΔE21) at D15 relative to D0. There is a markedly higher level of Xist induction in wt compared to KO clones. D IGV browser view of KDM6A profiles and peaks at D2, D4 and D7 of differentiation at the Xist locus in wt Tsix-stop cells. Merged data from two replicates of each time point are shown (see methods). KDM6A binding at the Xist gene promoter region and exon 1 is increased at D4 and lost at D7 following XCI. The scales of the profiles shown in (A) and (B) and (D) are indicated in the upper right corners
Chromatin analyses using Cut&Run with an antibody for KDM6A in wt Tsix-stop cells at D2, D4, and D7 show that KDM6A binding to the Xist promoter region increases at D4, when Xist expression levels continue to diverge between wt and Kdm6a KO cells, followed by a decrease at D7 (Figures 1C and 3D; Additional File 5: Figure S2A; Additional File 12: Table S6). A total of 582 KDM6A peaks mapped thoughout the entire genome in wt cells correspond to peaks of H3K27me3 enrichment, in contrast, in Kdm6a KO cells there is an increase in the number of H3K27me3 peaks that correspond to KDM6A peaks (870). This was observed both at D4 and D7, with a general decrease of KDM6A at D7 (Additional File 13: Figure S7A, B). Together, these results are consistent with KDM6A’s role in regulating H3K27me3 during cellular differentiation, such as in XCI, and with a genome-wide effect of Kdm6a KO on H3K27me3 levels (Additional File 13: Figure S7A, B). As expected, we observed KDM6A binding at promoters of known target genes with decreased expression in KO cells (Additional File 3: Table S3 and Additional File 13: Figure S7C) [14]. We next investigated the dynamics of KDM6A during early differentiation by qRT-PCR and protein blots, which showed a slight increase of both expression and protein levels during the early stages of differentiation followed by a decrease at D7 after XCI has occurred (Additional File 13: Figure S7D).
Allelic effects of Kdm6a KO on H3K27me3 enrichment were investigated in Tsix-stop cells in which the Xi is from strain 129 due to skewed XCI [3]. At D0 few differences between alleles and between wt and Kdm6a KO are seen (Figure 4A). However, the future Xi (maternal 129 X) shows enrichment in H3K27me3 along Xist, which is not seen on the future Xa (paternal cast X) (Figure 4A, B). This is evident both in wt and KO cells at D0, and is consistent with previous studies demonstrating maternal-specific H3K27me3 enrichment at Xist [26]. At D15, wt cells show high overall H3K27me3 levels on the 129 Xi compared to the cast Xa, as expected due to the onset of XCI (Figure 4A; Additional File 14: Table S7). In Kdm6a KO cells there is both an increase and a decrease in H3K27me3, the latter being specifically associated with a number of genes that gained expression due to the lack of Xist expression and partial failure of XCI (Fig. 2B; Additional File 15: Figure S8). Overall, the gain in H3K27me3 on the 129 Xi in KO cells was modest. We used the wt sample as a background to call Xi peaks that appear in the KO sample. There were only 50 Xi-specific peaks gained in the KO cells that were absent in the wt, with the majority appearing either in gene bodies or at the Xist gene. As a result, only an additional 0.014% of the Xi was associated with an H3K27me3 peak. Interestingly, only the maternal allele of Xist shows persistence of higher levels of H3K27me3 following Kdm6a KO, which would explain decreased Xist expression and impaired XCI (Figures 4B; Additional File 14: Table S7). The observed genome-wide increase in H3K27me3 probably contributes to the large number of autosomal genes with decreased expression in Kdm6a KO cells versus wt (Figure 1B; Additional File 1: Figure S1B).
Fig. 4
Kdm6a KO leads to increased H3K27me3 at Xist specifically on the Xi. A IGV browser views of gene expression (exp) and H3K27me3 peaks across the 129 and cast X chromosomes obtained by allelic analysis in Tsix-stop wt and KO cells (Tsix-Kdm6aΔEΔE17) at D0 and D15. The 129 profile is in pink (wt) or purple (KO) and the cast in light blue (wt) or dark blue (KO). XCI is skewed in Tsix-stop cells so that the 129 X will become the Xi and the cast X the Xa. The number of peaks for each condition and time point are in parentheses. There is a marked increase in Xist expression from the Xi in wt cells but not in KO cells at D15. H3K27me3 levels increase following differentiation on the Xi in wt and Kdm6a KO cells. B Same analysis as in (A), but zoomed in on the Xist gene. Allelic RNA-seq tracks of gene expression show very low expression from the 129 Xi allele in KO cells. A correlated Xi-specific increase in H3K27me3 enrichment at the Xist promoter is seen as an increase in peaks. No expression of Xist is observed from the cast allele, which is the Xa. The scales of the profiles are indicated in the upper right corners
We conclude that prior to the onset of XCI, Kdm6a KO has a minimal effect on H3K27me3 levels at Xist. However, following differentiation persistence of high H3K27me3 levels would explain low levels of Xist expression. Although we cannot fully exclude indirect effects of Kdm6a KO on Xist regulation, our data show association of KDM6A with the 5’ end of Xist during differentiation, suggesting that KDM6A may directly facilitate Xist expression during the female specific process of XCI.
留言 (0)