
記住我
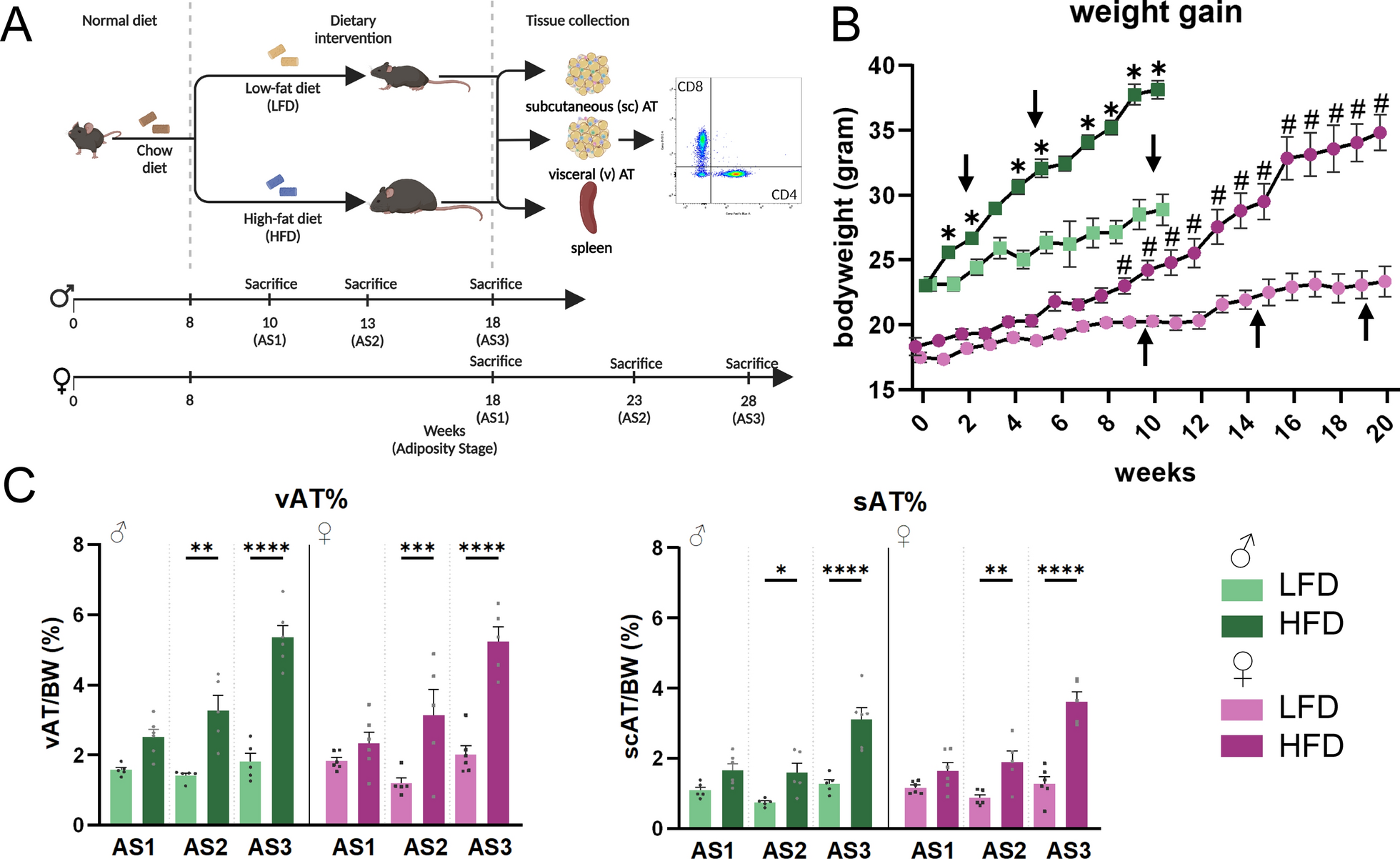


Literature shows that weight gain in female mice occurs later than in male mice, therefore, this was taken into account for our study design (Fig. 1A) [24,25,26]. We confirmed that body weight in HFD female mice only statistically increased after 9 weeks of HFD, while male mice show an increase after 1 week (Fig. 1B). To control for these differences in adiposity, male mice were sacrificed at the age of 10, 13, and 18 weeks. The start of sacrificing female mice started 8 weeks after the male mice: 18, 23, and 28 weeks (Fig. 1A, B). At these selected time points, we found that adiposity, defined as weight of the fat depot normalized to total body weight on the day of sacrifice, was not statistically different between male and female mice (Fig. 1C). Interaction analysis indeed revealed that sex did not affect vAT or scAT percentage, or the effect of diet or time (Supplementary File 1). Thus, these time points represent similar adiposity stages (AS) between male (10 weeks: AS1, 13 weeks: AS2 and 18 weeks: AS3) and female (18 weeks: AS1, 23 weeks: AS2, and 28 weeks: AS3) mice.
Taken together, our experimental setup allows us to compare male and female mice based on their adiposity, rather than the time of diet, and to leave the AS variable out of our statistical analyses (sex X diet interactions are summarized in Table 1).
The number of adaptive immune cells in AT depends on sex and the ASFirst, we looked at changes in the adaptive immune system in response to DIO (Fig. 2). In vAT, frequencies of CD19+ and CD4+ cells were increased after HFD independently of sex. In scAT, the number of immune cells was generally lower, but sex also impacted how immune cell populations changed over time.
Fig. 2
Sex affects the main adaptive immune cell regulation in vAT and scAT differently. A CD19+ B cells increase in the vAT and scAT of male and female mice with HFD, but the effect is more pronounced in the scAT. B CD3+CD4+ Th cells increase due to DIO in male mice with AS in vAT, but not in scAT. In female mice the number increases in the LFD group in vAT and diminishes the difference due to the diet. In the scAT only at AS3 CD3+CD4+ cells are upregulated in female mice. C CD3+CD8+ T cells are higher in female mice, but only in the vAT trends of an effect of the diet can be seen. (vAT: n = 6/♂ AS3 HFD, ♀ AS1 HFD, LFD; n = 5/♂ AS1 HFD, AS2 HFD, LFD, AS3 LFD, ♀ AS2 HFD, AS2 LFD (CD19, CD4), AS3 HFD, LFD (CD4, CD8); n = 4/♀ AS2 LFD CD8, AS3 LFD CD19; n = 3/♂ AS1 LFD; scAT: n = 6/♀ AS1 HFD; n = 5/♂ AS1 HFD, LFD (CD4, CD8), AS2 HFD, LFD, AS3 HFD, LFD (CD4), ♀ AS1 LFD, AS2 HFD, LFD, AS3 HFD, LFD; n = 4/♀ AS1 LFD CD19, AS3 LFD CD19, CD8;); p ≤ 0.05*, p ≤ 0.005**, p ≤ 0.001***
When performing quantification of the different lymphocyte compartments in response to HFD, we identified that in the vAT, the count of CD19+ cells increases with adiposity in HFD independent of sex, but it happens at an earlier AS in female mice. In the scAT we see a similar pattern (Fig. 2A). In the vAT, we detected an increased number of Th cells (CD3+CD4+) in female mice at AS1 and in male mice at AS3 due to the HFD. In the scAT, lower numbers of CD4+ T cells were detected, but we did identify an accumulation of CD4+ T cells in female mice on HFD (Fig. 2B). In the CD8+ T cell compartment (Fig. 2C), we identified a high inter-subject variation of CD8+ T cells in the vAT after HFD in female mice. Due to this, no significant differences were identified. In the scAT, we could detect an effect of diet, but looking at both sexes separately, we did not detect a change in CD8+ numbers.
Within the different Th cell subsets (Fig. 3), DIO did not affect Th1 (CXCR3+CCR6−) cell numbers in vAT. However, in scAT a clear increase of Th1 cells at AS3 in female mice on HFD was found (Fig. 3A). Importantly, the overall effect of HFD on Th1 levels appeared independent of sex.
Fig. 3
Pro-inflammatory Th cell subsets are differently regulated in male and female mice in AT. A Pro-inflammatory Th1 (CXCR3+CCR6−) B Th17 (CCR6+CXCR3−) and C Th17/1 (CXCR3+CCR6+) cell counts in visceral and subuctaneous fat depots of male and female mice. D Treg (CD25+) counts in the AT depots. (vAT: n = 6/♂ AS3 HFD, ♀ AS1 HFD (Th17, TH17/1, Treg), LFD; n = 5/♂ AS1 HFD, AS2 HFD, LFD, AS3 LFD, ♀ AS2 HFD, AS2 LFD (Th17), AS3 HFD, LFD); n = 4/♀ AS2 LFD Th1; n = 3/♂ AS1 LFD; scAT: all groups n = 5, only ♀ AS1 HFD n = 6); p ≤ 0.05*, p ≤ 0.005**, p ≤ 0.001***
As seen in Fig. 3B, changes in Th17 (CCR6+CXCR3−) cells due to the diet differed between male and female mice based on AS (Fig. 3B). In female mice Th17 are more present at AS1 in vAT and in male mice only a trend of upregulation due to HFD at AS3 was seen. Furthermore, the higher number of Th17 cells at AS1 in HFD female mice in vAT and scAT subsides with a higher AS. In the scAT, a similar trend was found for the effect of sex on the diet response.
For Th1/17 cells (CCR6+CXCR3+), we identified that sex affects how mice react to DIO in different AS stages in vAT. The number of Th1/17 cells at AS1 was very low in male mice, but with increasing adiposity, they tended to accumulate in the HFD group (Fig. 3C). In females, Th1/17 accumulated at and early AS and were significantly elevated at AS2. However, this difference subsided in AS3, due to an increase in LFD mice. In the scAT the amount of these cells was very low, although they tended to accumulate in this depot over time. While multiple comparisons did not identify a significant difference between the dietary interventions, a simple effect of diet was observed when combining the data of male and female mice from all AS. Overall, in female mice, the upregulation of pro-inflammatory Th cell subsets was diminished with increasing AS. In the scAT, an upregulation was only identified in female HFD mice.
Regulatory T cells (Tregs, CD25+) did not show an interaction with sex, even though especially in male mice clear changes in Treg numbers were observed due to HFD (Fig. 3D). Pairwise comparisons reveal that at AS2 Tregs were increased in female mice on HFD. In male mice this occurred at AS3, while in female mice Tregs subsided to baseline level. In scAT we found a strong overall effect of diet. At all adiposity stages a trend toward a higher Treg count in the HFD fed female mice was observed, which was significant at AS3. In the male mice, only at AS3, Tregs tended to be increased by DIO.
Taken together, CD19+ cells were increased in the DIO model of both sexes, but at an earlier AS in female mice. In the T cell compartments, the number of cells was also increased at AS1 and AS2 in female mice, while at AS3 no differences were observed due to an increase in T cells in control-diet fed animals. In contrast, male mice displayed a steady increase. In the scAT only CD4+ T cells displayed changes between different AS. To reveal whether the observed changes in the immune compartments were specific to AT in these mice, we also analysed the same cell populations in the spleens (Fig. S2). Overall, no clear skewing to pro-inflammatory cells due to HFD was detected. Furthermore, the identified differences were not in line with the changes in the fat depots, showing that the described changes in the white fat depots were not due to general changes in a primary lymphoid organ, and therefore occur independently.
HFD upregulates myeloid cells in different fat depots based on sexThe main immune cell populations in the AT are myeloid cells (CD11b+) [7, 27], therefore we assessed their overall presence in the AT depots (Fig. 4). Interestingly, sex strongly affected how diet changed this cell compartment at different AS in vAT. Myeloid cells were increased with adiposity in male mice, while they remained stable in the LFD group. In female mice, myeloid cell numbers were higher in the HFD group at AS1, but with increased adiposity these levels decreased and the numbers in the LFD group increased. In contrast, in scAT, the amount of CD11b+ cells was unaffected in male mice, while HFD lead to a stronger upregulation in female mice, reflecting a clear effect of sex on the diet response (SEX x DIET: F(2,50) = 7.03, p = 0.011; Table 1). In short, in male mice dietary interventions impact myeloid cells in the vAT, but in females this effect is pronounced in the scAT.
Fig. 4
Myeloid cells (CD11b+) are upregulated in different AT depots depending on sex. A CD11b+ cells are increasing with AS in male mice, while they are decreasing in female mice. B In male mice CD11b+ cells are not affected by DIO. But in female mice DIO upregulates them at all AS. (n = 6/♂ AS1 HFD scAT, AS3 HFD vAT, ♀ AS1 HFD, LFD vAT; n = 5/♂ AS1 HFD vAT, AS2 HFD, LFD scAT, AS3 HFD scAT, LFD vAT ♀ AS1 LFD scAT, AS2 HFD, LFD, AS3 HFD, LFD; n = 4/♂ AS2 LFD vAT, AS3 LFD scAT; n = 3/♂ AS1 LFD vAT); p ≤ 0.05*, p ≤ 0.005**, p ≤ 0.001***
Sex differences in AT depots are independent of time on diet and ageInterestingly, AS3 in male mice and AS1 in female mice had similar patterns of immune cell accumulation in the AT depots. Since these groups differ in AS but reflected the same diet duration and age (Fig. 1), we compared these groups directly to each other (Fig. S3). Although some immune cell populations are not significantly different between male and female mice after 10 weeks, myeloid cells and Tregs in the vAT were only increased in male mice. This comparison shows that even at the same age and after the same dietary intervention period, some sex differences are apparent.
Only in male mice, HFD leads to inflammasome activation in myeloid cells of vATNext, we investigated inflammasome activation in the different cell compartments. In male mice, DIO led to inflammasome activation in vAT myeloid cells. On the contrary, diet had no effect on inflammasome activation in female mice. However, the baseline % of cells with an activated inflammasome was always higher in female mice if compared to male mice. In addition, sex strongly affected inflammasome activation due to diet and AS. In the scAT the differences were smaller, but overall, a similar trend can be seen (Fig. 5A).
Fig. 5
Only in male mice, CD11b+ cells inflammasome activation is upregulated with DIO. A Myeloid cells (CD11b+) have an upregulated inflammasome activation due to DIO in male mice. In Th cells (B, CD3+CD4+), B cells (C, CD19+) and CD8+ T cells (D) changes in inflammasome activation are not clearly linked to adiposity. (n = 3–6/group, see Figs. 2 + 4); p ≤ 0.05*, p ≤ 0.005**, p ≤ 0.001***
In cells of the adaptive immune system of male and female mice, we found a trend towards higher inflammasome activation at the lowest AS in both sexes on HFD, but at higher adiposity, no clear inflammasome activation in AT could be identified (Fig. 5B–D). Similar findings were observed in the spleens of these mice (Fig. S4). Together, this suggests that inflammasome activation is male-specific in DIO and most prominent in the vAT.
Systemic inflammatory cytokines mimic inflammation in the vATTo understand if the differences in the AT depots correlated with systemic inflammatory changes, we assessed pro-inflammatory cytokines in the blood plasma. Since HFD leads to low grade chronic inflammation, with an upregulation of several different cytokines, we computed an inflammation z-score (including IL-23, IL-1α, IFN-γ, TNF, MCP-1, IL-1β, GM-CSF, IL-17A) to get an overview of the inflammatory state (Fig. 6). Only simple effects of the different ASs could be observed, but no effect of the dietary intervention or an effect of sex on DIO. There is a trend that the z-score increases for male mice on HFD depending on AS. In contrast, the same seems to be the case for female mice on LFD. However, from the chosen cytokines, not all were detectable in the blood plasma of each mouse, leading to a high variation in different groups.
Fig. 6
Pro-inflammatory cytokines in the blood plasma mirror the vAT inflammatory stage. Based on a z-score of pro-inflammatory cytokines (IL-23, IL-1α, IFN-γ, TNF- α, MCP-1, IL-1β, IL-17A, GM-CSF), no significant change in systemic inflammation due to adiposity can be found. (n = 6/♂ AS1 HFD, AS3 HFD ♀ AS1 HFD, LFD, AS3 LFD; n = 5/♂ AS1 LFD, AS2 HFD,♀ AS2 HFD, LFD, AS3 HFD; n = 4 ♂ AS2 LFD, AS3 LFD)
Given its direct correlation with inflammasome activation, we additionally assessed IL-1β levels separately from the compiled z-score. In line with the ASC specking data (Fig. 5), HFD increases IL-1β levels in plasma of male mice, although this increase did not reach statistical significance (Fig. S5). In female mice, however, there was an increase in IL-1β plasma levels due to HFD at AS1 and AS2, which is in contrast to the overall finding of ASC-specking in AT-derived immune cells. At AS3, no effect of diet could be observed in female mice, which is in line with ASC-specking.
留言 (0)