
記住我
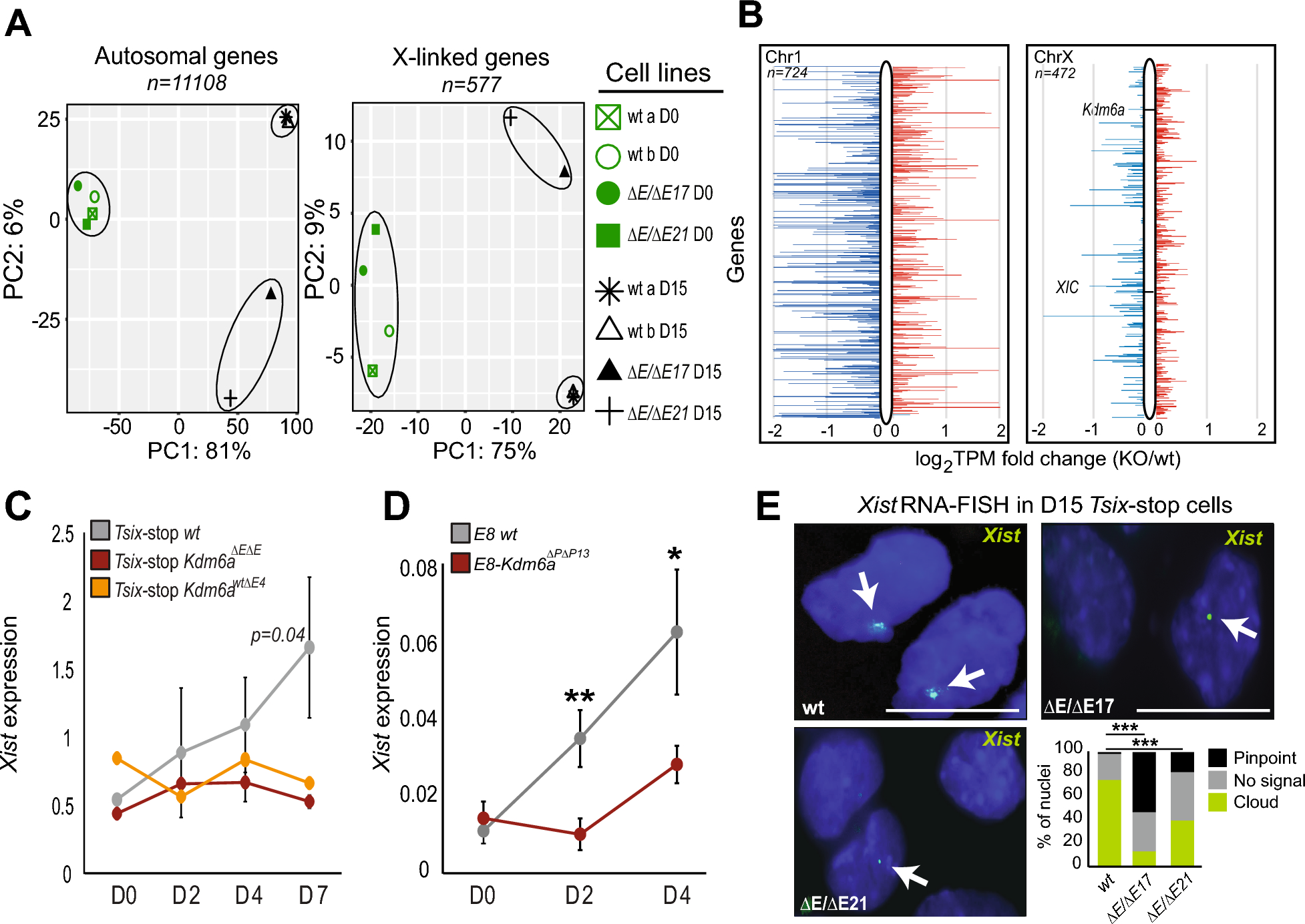

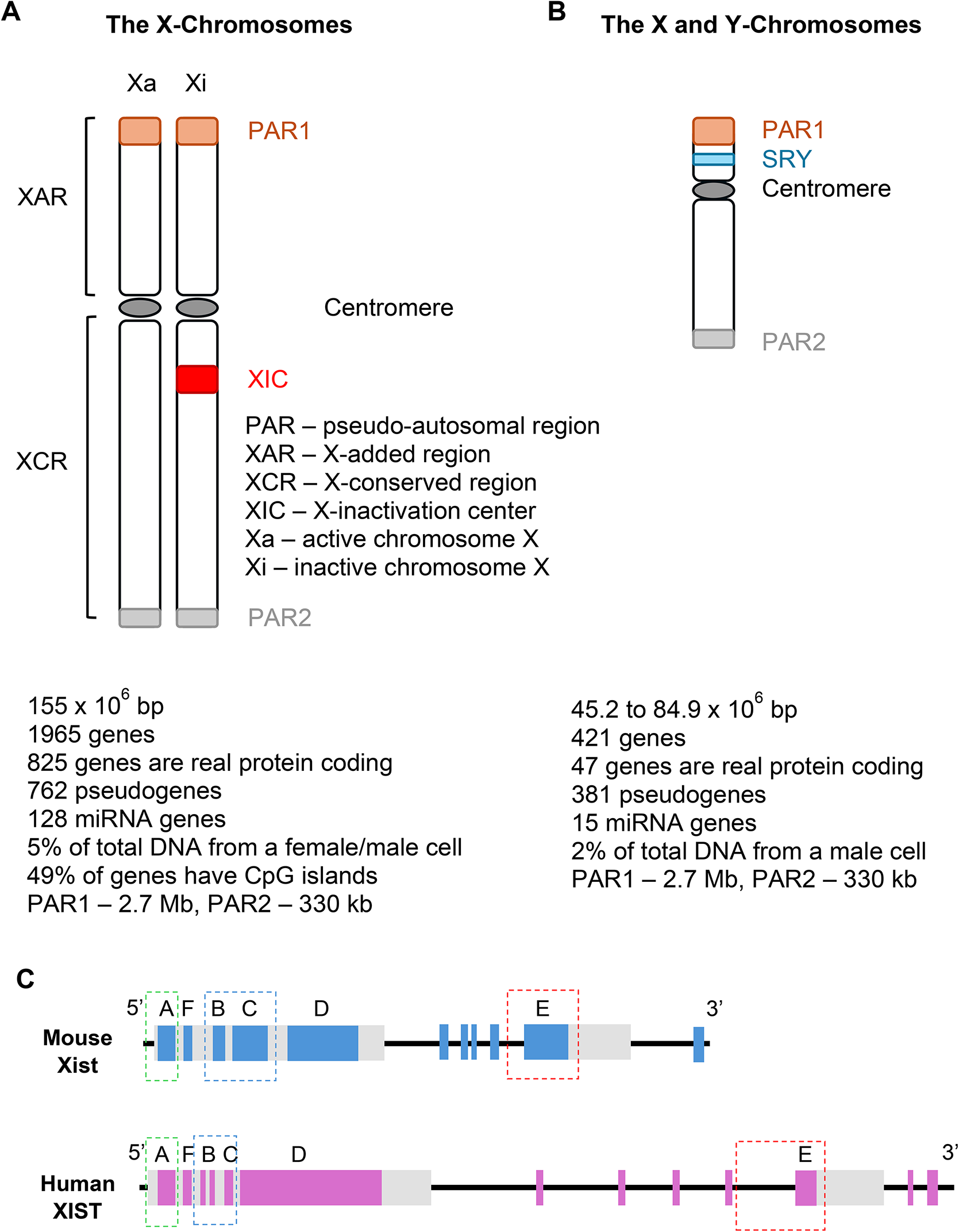

The XX and XY, the sex chromosome complements in humans. In eukaryotic cells, there are two X chromosomes, an X-inactive (Xi) and an X-active (Xa) in female cells, and an Xa and an Y chromosome in male cells. The Xi requires: (i) the establishment of specific post-translational modifications of histones, (ii) the creation of specific DNA methylation patterns of X-linked gene promoters, and (iii) changes in the higher-order of chromosome [27, 28]. Under evolutionary pressure, this sexual disparity in the number of X chromosomes controls both the sex-specific gene expression and the crosstalk between sex chromosomes and autosomes [29]. This is the genetic base of human sexual determinism. It is worth mentioning that even though the X chromosome accounts for 5% of active genes in humans and has a sizable contribution, it is considered as a chromosome with a low density of genes [30]. The X chromosome, with ~ 155 mega bases (mb) is 2.6-fold the size of the Y chromosome, and is “gene-rich” versus the Y chromosome, with greater than 1000 genes and 700 pseudogenes, while the Y chromosome is ~ 60 mb, is “gene-poor”, having less than 140 genes, Fig. 1, A, B [31, 32].
This numeric imbalance between the number of sex chromosomes between males and females, known as sex chromosome dosage inequality, is corrected in the first days of embryonic development by dosage compensation, a mechanisms meant to equalize sex chromosome-linked gene expression before gonadal differentiation [33]. Dosage compensation is achieved in a two-fold manner in mammals, first, by inactivation of one of the two X chromosomes in females, and second, by upregulation of X-linked genes, to balance the expression levels between X-linked and autosomal genes [34]. X chromosome inactivation is largely mediated by the lncRNA XIST, which acts to recruit repressive complexes for silencing one X chromosome. However, despite being on the Xi, about 20% of X-linked genes (termed escapees) escape from XCI and remain active. Some of these escapees have an Y chromosome paralog, and thus, they have equal expression between sexes. Other escapees are expressed exclusively from the X chromosome and will exhibit higher expression in females. Some escapees have been directly correlated to specific disease characteristics [35,36,37]. A less elucidated mechanism for dosage compensation to maintain a balance between X-linked and autosomal gene expression is doubling the transcription from the Xa chromosome [34].
Fig. 1
Significant features of the X and Y chromosomes. A. The female signature consists of two X chromosomes, the Xa and Xi pair. B. The Xa and one chromosome Y represent the male mark. C. The XIST RNA molecule repeats between human, and mouse XIST/Xist are conserved. XIST/Xist RNA contains six types of repeats, labeled A-F. The E-repeat is located at the 5 end of exon 6 of XIST/Xist and at the 5 end of exon 7 of XIST/Xist, respectively; all other repeats are contained within the large first exon of XIST/Xist. The approximate copy number and monomer length for each repeat block is: A repeat (mouse 8x ~ 50 nt; human 9x ~ 100 nt); F repeat (mouse 2x ~ 16 nt; human 2x ~ 16 nt); B repeat (mouse 32x ~ 7 nt; human 29x ~ 7 nt); C repeat (mouse 14x ~ 115 nt; human 1x ~ 115 nt); D repeat (mouse 10x ~ 290 nt; human 26x ~ 290 nt); E repeat (mouse 50x ~ 25 nt; human 28x ~ 25 nt). The dashed boxes indicate the repeats that contribute to gene silencing (green), Polycomb Group (PcG) protein recruitment (Blue) and XIST/Xist localization (red)
The Good of XIST relates to the lyonization of gene expression. During dosage compensation, one of the X chromosomes in female cells is transcriptionally silenced at random, in the space of the X inactivation center (XIC) This phenomenon is also known as “lyonization’ in honor of Mary Frances Lyon, its first describer [38]. Lyonization starts with the upregulation of the lncRNA XIST from one allele in each cell [39,40,41]. XIC is about 1 Mb region of X chromosome, at Xq13.2 and contains some protein-coding genes including, Rnf12, as well as several lncRNAs including RepA, Tsix, Xite, Jpx, Ftx, and Tsix [28]. Whereas there is agreement that RNF12 is one of the proteins form XIC which by targeting REX1 is directly involved in XIST activation [42, 43], there are other proteins involved and the reader is referred to a recent review for an in dept analysis of XIST interaction with different proteins [44].
Xist/XIST, a 15 kb lncRNA in mice and 17 kb lncRNA in humans, is transcribed only from the Xi chromosome, and even if poorly conserved, XIST/Xist contributes and is required for XIC/Xic. Currently, because of the concept of XIST being expressed from only the Xi chromosome, it is considered a typically female exclusive transcript, and thus, not expressed in males [45]. Under normal conditions, detection of XIST presence in males was difficult, if not impossible, considering the low number of XIST molecules per cell and the analytical resolution of the in situ hybridization methodology, in the range of 100–200 kb as determined by the probe size, prevalently used for its detection [46]. The use of higher resolution qPCR and RNA sequencing significantly improved XIST detection over in situ hybridization and fluorescence [47]. XIST expression is associated with the X chromosome from which it is produced, is acting only in cis, and is a molecule created of several conserved tandem repeats labeled A to F [48]. The XIST repeats, five major structural building blocks - A, F, B/C/D, E, and exon 6/7, Fig. 1C, described for mice are conserved in their human counterpart [49, 50]. Deletion analyses of mouse Xist repeats revealed the synergistic and coordinated functions of these modules, established that XIST/Xist is a multitasking lncRNA, and helped to find out the specific RNA Binding Proteins (RBPs) that interact with the functional domains, and finally, confirmed their involvement in Xic from its beginning till at the end [51,52,53]. Exon 6 is considered a poor protein binding domain. A protein that is reported and confirmed to bind to the center of the Exon 6 is hnRNPU, while several proteins like: chromodomain-helicase-DNA-binding protein 4 (CHD4), enhancer of zeste polycomb repressive complex 2 subunit (EZH2), SUZ12 Polycomb Repressive Complex 2 Subunit (SUZ12), and polycomb group protein CBX (pCBX) associate with the sides of the Exon 6 [50]. Considering this distribution, a poor protein binding center domain and sides-enriched with binding proteins, it has been speculated that such scattering may help to bring its two ends to proximity, which is concordant with the spatial structure of Exon 6. Binding the hnRNPU proteins along with the BCD, a major structural domain of Xist which also binds the CDKN1A Interacting Zinc Finger Protein 1 (CIZ1), makes Exon 6 a participant in tethering Xist to Xi. Also, like the domain BCD, the compact folding of Exon 6 excludes the m6A methylase complex and, by this, contributes to the specificity of m6A modifications [50]. Thus, according to the current evidence, Exon 6 participates in three processes related to Xist structure/activity.
Structural modularity of XIST/Xist RNA protein complexesRepeat A, the focal point of XIST/Xist silencing, is highly conserved in mammals and has been shown to be indispensable for gene-silencing [54]. Repeat A essentiality for transcriptional silencing was confirmed by proteomic methodologies which mapped its RNA–protein interactions [55]. This repeat is the focal point of initiation of transcriptional silencing by recruiting SPEN (SPlit Ends), which contains the repressive domain SPOC (SPEN Paralogue/Orthologue C-terminal) that recruits the repressive factors NCoR/SMRT (Nuclear receptor Co-Repressor/Silencing Mediator of Retinoic acid and Thyroid hormone receptor) complex. After recruitment of the NCoR/SMRT complex, one of the first events that takes place is the deacetylation of histone H3 and H4. These are the first steps - recruiting and deacetylation - in initiating gene silencing. Given the biological importance of the A domain, repeated efforts have been made to elucidate its spatial structure to associate it with XIST biological function. Currently, the prevailing view is that repeat A functions as a nucleation center for dynamic protein assembly which organizes them as a hub of transcriptional silencing [56]. Deletion of repeat A abolishes gene silencing, which also leads to a defect in XIST/Xist local spreading and spatial exclusion of active genes from the XIST/Xist-coated nuclear compartment [57, 58]. Thus, the Repeat A domain contains RBPs involved in transcriptional silencing, splicing regulation, DNA methylation, m6A modification, and nuclear lamina attachment [50].
Repeats B and C, the heart of Polycomb Repressive Complexes mobilization, are important for locking the silent state of the Xi, through the recruitment of the PcG protein complexes to the Xi after the initial establishment of XIC by the repeat A module. Even if widely scrutinized, the mechanisms and sequences of events involved in PcG recruitment, are still an area of hot investigation [59]. As the two Repeats B and C are cytosine-reach domains, they serve as a docking site for the Heterogeneous Nuclear Ribo Nucleo Protein K (hnRNPK), which by binding the PcG Ring Finger 3/PcG Ring Finger - Protein Regulator of Cytokinesis 1 (PCGF3/PCGF5–PRC1) complex decorates the chromatin with the histone H2AK1219 mono-ubiquitination mark (H2AK119ub), which is the primary signal for recruiting the Polycomb Repressive Complex 2 (PRC2), via its recognition by Jumonji- and AT-rich interaction domain (ARID)-domain-containing protein 2 (JARID2) from the PRC2 [51]. Both repeats are binding to the hnRNPK, but it seems that the repeat C is involved in XIST localization via its interactions with the heterogenous nuclear ribonucleoprotein U/scaffold attachment factor A (hnRNPU/Saf-A) and Yin Yang 1 (YY1) [60]. While polycomb recruitment is of the essence for chromosome-wide silencing, the contribution of PRC1 and PRC2 remains to be established. So far, repeat B is a prerequisite for XIST RNA to spread and by doing this, is an active participant in gene silencing. Moreover, the deletion of repeat B or of hnRNPK showed that the recruitment of PRC1 and PRC2 is compromised. To conclude, the domains B/C are key players in heterochromatin formation and topological reconfiguration of the X chromosome and thus, in long lasting transcriptional silencing lengthwise of the Xi [59].
Repeat D, the neglected child of Xist/XIST, was described initially as an ~ 200 bp motif [40] and later shown to contain 10 truncated Repeat D motifs located at nucleotides ~ 5200–7900 toward the 3’ end of exon 1. It was reported to be the most complex of the Xist/XIST repeats [61]. Deletional analysis did not show any changes in phenotypes related to Xist/XIST coating or PCG recruitment in mouse embryonic fibroblasts [51]. Given the lack of functional data of the structurally complex repeat D, it has been suggested that the repeat might have a role in the early stages of XCI [59]. By comparison to the mouse, the human Repeat D is larger, suggesting that it may replace the function of repeat C, which in humans is reduced to one incomplete motif [59].
Repeat E, the hub of Xist/XIST localization to Xi, is important for tethering Xist/XIST by interacting with several RBPs necessary for anchoring to the Xi territory. Different methods and methodologies used to map the RBN complexes associated with each domain, established that the domain E which binds CIZ1 and hnRNPU proteins, attaches the Xist/XIST RNA to the Xi [62,63,64]. CIZ1 is one member of a group of matrix-associated proteins shown to interact with Xist/XIST and strongly enriched at the site of Xist/XIST RNA, in both human and mouse models [62]. Additionally, it was proposed that repeat E may participate in the recruitment of PRC2, KMT5A, and CIZ1, all of which operate independently of silencing or chromatin remodeling [65]. Furthermore, the binding of polypyrimidine Tract Binding Protein 1 (PTBP1), matrin-3 (MATR3), CUGBP (CUG triplet repeat RNA binding protein) Elav-Like Family Member 1 (CELF1), and Transactive Response DNA-binding protein 43 (TDP-43) creates a distinct functional complex which stabilizes Xist/XIST coating after the initial wave of transcriptional silencing and PcG recruitment by repeat A and repeats B/C. It is worth mentioning that, even if the polypyrimidine tract binding protein 1 (PTBP1), with TAR DNA binding protein (TARDBP), ADP-ribosylation factor 6 ( ANF622), and serine and arginine rich splicing factor 7 (SRSF7) are spatially segregated on the E domain, their functionality remains to be demonstrated [53]. By doing this, repeat E plays a central role in the maintenance of XCI likely, via the assembly of a specialized phase-separated sub-nuclear compartment necessary for the efficient maintenance of XCI [53].
Repeat F, the shortest of all domains, comprises two copies of 10 bp in mouse Xist RNA. As the deletion studies have produced conflicting results regarding its role in XCI, more work is needed to reconcile its possible roles, either as a binding site for YY 1, and/or as a participant in transcriptional silencing via the loss of tethering of Xi to the nuclear lamina [56, 66]. Recent studies reported that the Lamin B receptor binds to three sites across Xist, with the most prominent encompassing the entire F repeat, and tethers the Xi chromosome to the nuclear lamina, a region where gene expression is silenced [56]. Nuclear lamina is required for reshaping the chromatin structure to allow its proficient association with the transcriptionally active genes and silencing [67,68,69].
Xist/XIST positive-negative regulatorsThe genes of three lncRNAs TSIX, XITE and LINX, whose main role is to randomly inactivate one X chromosome in the embryo development reside within the same topologically associated domain (TAD) from XIC. On the same genomic space, the Xist/XIST promoter shares TAD with the other three lncRNAs positive regulators, JPX, FTX, XERT, and the protein-Rnfl12 [70]. Outside of this brief outline of XIC, it should be mentioned that other parts within the XIST locus have been reported to contribute to XIST upregulation [71]. Work related to SPEN, essential for gene silencing during XCI, revealed a cis-acting positive feedback mechanism that boosts XIST expression by either mutual repression when XIST silences one or some of its cis-repressors, or by mutual activation, by XIST induction of cis-activators [72, 73]. The studies to understand the who and how different players involved are working together have established that the region encoding for lncRNA XIST and TSIX, the antisense transcript of Xist/XIST that can mediate its repression, are essential for initiation, establishment, and propagation of XIC [74]. There is agreement in claiming that Tsix is the one that dictates which chromosome X will be Xa by modifying the chromatin state of that chromosome, and by managing the DNA methylation of the Xist/XIST promoter [75]. However, the gene expression from corner to corner of the Xi chromosome does not happen at the pseudo-autosomal regions (PARs) that are present on both X and Y chromosomes, Fig. 1A, B [32]. On the PARs, the repressive events that mark the inactivated chromosome: (i) exclusion of RNA polymerase II, (ii) DNA methylation at promoters and (iii) adding in place histone modifications, that are missing from the PARs genes which are not imprinted [76]. As illustrated in Fig. 1A, B, in humans, PAR1 is of 2.7 Mb, while PAR2 is of only 330 kb [77]. The expression of genes from the PARs is significant because PAR1 genes are highly expressed in males, 14 out of 15 genes, making their expression to surpass the combined Xa and Xi expression in females [78].
Xist/XIST RNA – a master regulator of XCIXist/XIST associates with 81 unique binding proteins to form different ribonucleoprotein complexes, more than 20 through direct RNA-protein interaction and others through indirect protein-protein interaction [55]. While different proteins have been described as participating in XIC, recruitment of Polycomb group proteins following Xist/XIST RNA coating occurs in the early stages of Xi establishment. Polycomb complexes PCRC1 and PCRC2 are primarily in charge of histone modifications that are paramount for XIC. While the PRC1 complex catalyzes ubiquitination of histone H2A at lysine 119 (H2AK119u1) [79], the PRC2 complex catalyzes lysine methylation -H3K27me3 [80]. Recent studies show that the non-canonical PcG RING finger 3/5 (PCG3/5)-PRC1 complex initiates recruitment of both PRC1 and PRC2 in response to Xist/XIST expression, highlighting the fact that the mechanisms of XIST-mediated recruitment of PcG proteins are far from being fully understood [52, 81].
Before super-resolution structural studies, it was reasoned that Xist/XIST together with its effectors proteins, other lncRNAs, and chromatin-induced modifications, accumulate over the whole chromosome and by their distribution from corner to corner, control the expression of all genes from X chromosome [82]. Super-resolution microscopy, however, has proved that XIST distributes on Xi as 50–150 nm diffraction-limited foci in differentiated cells [83]. This new concept of Xi being carried out by protein foci, despite being revolutionary, cannot explain how the known effector proteins interacting with Xist/XIST are capable of silencing more than 800 genes spanning 150 million base pairs. Nevertheless, super-resolution microscopy combined with single-particle tracking provided solid data showing that Xist/XIST foci are locally confined and once in place, they stimulate transient dynamic protein compartments formation around a slowly exchanging Xist/XIST, referred to as supramolecular complexes (SMACs). In SMACs, the rapid binding and dissociation of most Xist/XIST-interacting proteins is the driving force behind local protein gradients stretched over broad regions of the X chromosome responsible for silencing [84].
However, the mechanisms behind other functions of Xist/XIST remain to be revealed. The female exclusive activation of XIC is mediated by a tightly controlled balance between X-encoded XIC activators and XIC repressors [85]. Initiation of XIC is stochastic, and a negative feedback loop involving rapid silencing of some X-encoded XIC activators can prevent the inactivation of the second X chromosome [86]. The stochastic model of XIC dynamics was confirmed, at least partially, by mathematic modeling and machine learning algorithms [87]. It is assumed that once XIC is established the Xi enters the maintenance stage which is watched over by several molecular mechanisms: (i) continuous XIST expression, (ii) histones hypoacetylation, (iii) DNA methylation, and (iv) acquisition of histone macroH2A and CULLIN3/SPOP ubiquitin E3 ligase in the late stages of Xi establishment. Once established, XIC is stably propagated, but it was shown that can be reversed in vivo and in vitro by pluripotent reprogramming and by DNA hypomethylation [88, 89]. The developmentally orchestrated process of XIC is considered perpetual for the lifespan of female cells and creates a rule according to which only one Xa chromosome is typically present in human cells. However, this genetic dogma does not apply to cancerous cells of both sexes when supernumerary X chromosomes are detected and XIST expression is dysregulated. While the “re-apparition” of XIST in somatic cells is well-documented for some forms of cancers, in several autoimmune and neurological diseases, its presence in females, and particularly in male PAH patients, is a novelty [90, 91].
The silenced state of the Xist/XIST geneWhile for females (both mice and humans) the long-noncoding RNA Xist/XIST is indispensable for initiating and maintaining XCI of one of X chromosomes during early development, the expression of XIST in male normal tissues is barely detected, but not zero. Xist/XIST presence was demonstrated by qPCR in the lung tissue of an experimental murine model of PAH, in the lung tissue of human PAH patients, and in human lung endothelial cells from lung PAH explants [92], as well as by sequencing in different human cancer tissues [93]. Significantly, XIST has also been detected in 0.6% of male somatic tissue adjacent to the tumors, presumed to be normal, and in 0.2% of normal male tissues. Hence, the values of XIST in normal somatic tissues are small, but they show that either XIST shutting off after the dosage compensation completion is not absolute, mainly for female cells, or that XIST gene can be reactivated in somatic cells, in both male and female, by mechanisms remaining to be elucidated. Even if the X chromosome from the male is Xa, under the circumstances described above or in various settings just starting to be identified, XIST can be detected in males’ somatic cells.
The Good of Xist/XIST is also found in sex-specific regulation of aging. Aging in living organisms, also known as senescence, is generally described as a cumulative, irreversible process resulting in decreased function and increased risk of death [94]. Aging is accompanied by alterations in metabolism, body composition, hormone status, and accumulation of abdominal fat. Studies of sex differences in longevity indicate a 4–10 years longer life in females versus males [95]. Using the Four Core Genotypes mouse model, Davis et al., have shown that chromosome X content has been linked to a longer life span and the presence of two chromosomes X contributes to increased longevity regardless of the hormonal status [96]. Recent studies demonstrate a female-specific feature of hypothalamic aging [97]. Using single nuclei sequencing of the aging female mouse (19–24 months) hypothalamus and bioinformatics, Hajdarovic and colleagues found that the major cell types of the hypothalamus undergo widespread transcriptional changes. The hypothalamus is a critical brain region for the regulation of physiological homeostasis, including feeding, metabolism, and sleep. The most upregulated genes included the female specific XIST and TSIX. Upregulation of Xist expression with age was not detected in male mice. Moreover, a learning machine approach indicated that X chromosome gene expression is sufficient to predict cellular age, and Xist is the second most important predictor of age.
A different potential explanation for the females longer life compared to males may be due to the suboptimal mitochondrial function in males and the antagonist pleiotropy of gene function between the sexes [94]. Both the mitochondrion genome and the X chromosome are asymmetrically inherited in mammals – in the sense that through evolution, the associated genes spend relatively more time under selection in females and thus, expected to be better optimized for function in females than in males and to contribute preferentially to the aging phenotype.
The Bad of XIST is found in the XIST driven pathologies. Many recent studies confirmed XIST participation in a substantial number of pathologies like neurological, cardiovascular, lung, kidney, pancreas, skin, autoimmune, bone diseases and cancers [91, 92, 98,99,100,101]. XIST as a nuclear regulatory RNA, not only dictates the process of dosage compensation by establishing and maintaining XCI, but also may influence gene expression by epigenetic management and as a transcriptional regulator, via RNA processing and chromatin organization. Dysregulation of its expression and defects in its functions can cause severe phenotypes and increase susceptibility to genetic disease.
XIST and autoimmune diseasesWhile females have an enhanced capability to solve infections when contrasted to males, they do have an increased vulnerability to developing autoimmune diseases [102]. Autoimmune diseases, occurring in 3–5% of the population are behind the heart disease and cancer, as the third most prevalent human disease category affecting females more than males; 4 out of 5 patients are female [
留言 (0)