
記住我
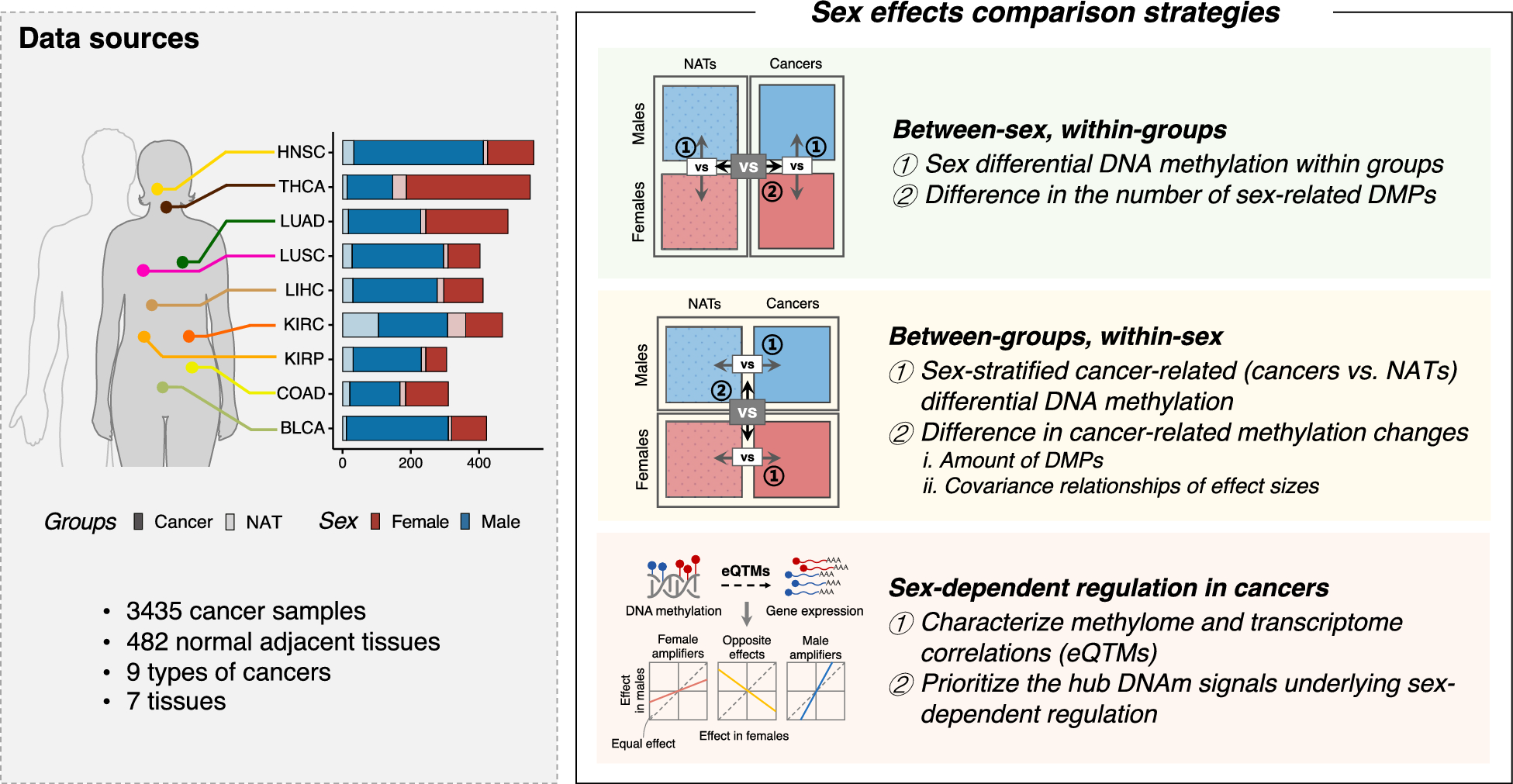


To investigate the sex effects on DNAm regulation in cancers, we obtained DNAm data and RNA-seq data with sufficient sample sizes (≥ 10 each sex per cancer) in nine major non-reproductive cancer types from TCGA (Supplementary Table 1). These cancer types include HNSC, THCA, LUAD, LUSC, LIHC, KIRC, KIRP, BLCA, and COAD.
The overall study design is summarized in Fig. 1. First, we systematically compared the extent of sex differential DNAm between the paired cancers and NATs to determine whether the observed sex differences are reorganized in cancers. Next, employing a sex-stratified case–control strategy, we examined the amount, correlation, and magnitude of cancer-related DNAm changes between sexes to decipher how sex effects influence the aberrant DNAm patterns in cancers. Finally, we characterized correlations between the methylome and transcriptome, manifesting as eQTMs, to prioritize hub signals underlying sex-biased DNAm regulation in cancers. Through this DNAm-focused integrative analysis, our findings enhance understanding of molecular regulation in sex effects on cancers.
Fig. 1
Study design. Schematic illustration of the key analyses used to investigate the impact of sex on DNAm in nine types of non-reproductive cancers. DNAm, DNA methylation; DMPs, differentially methylated positions
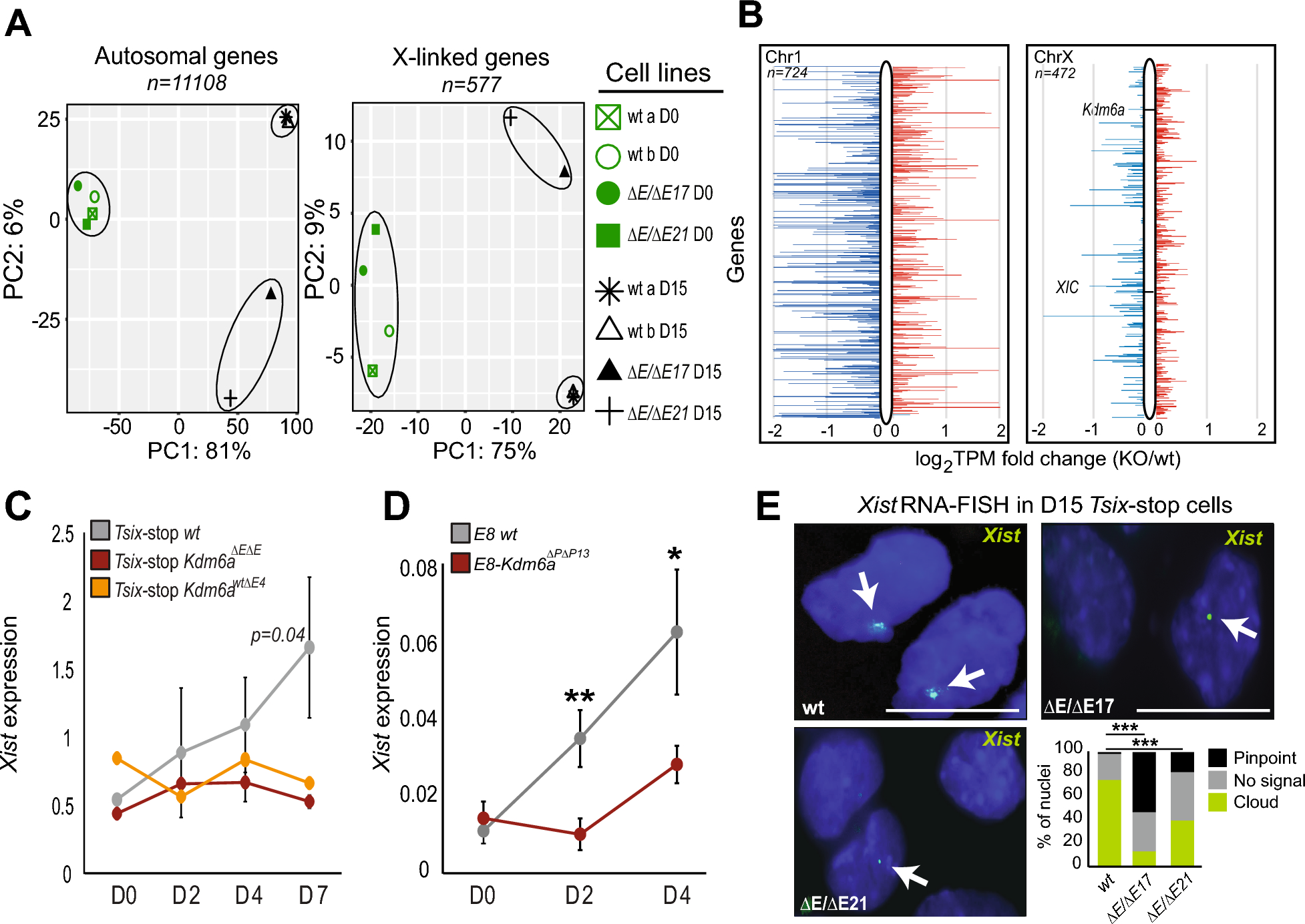
Attenuation of sex differences in DNAm across cancersTo determine whether sex-related differential DNAm is reorganized in cancers, we compared the number of sex-related differentially methylated positions (sDMPs) between the paired cancer and NAT samples across nine cancer types, using a conservative permutation-based method (Methods). First, we quantified sex bias effects for cancers and NATs separately. In NATs, we identified 8,833 non-redundant sDMPs, ranging from 3,211 in BLCA to 8,084 in KIRC (Bonferroni-adjusted P-value, Pbonf < 0.05, Supplementary Table 2). Meanwhile, 6,361 sDMPs were identified in cancers, ranging from 24 in BLCA to 5,968 in KIRC (Pbonf < 0.05, Supplementary Table 2). Then, by comparing the number of sDMPs, we observed a significant reduction in the number of sDMPs in cancers compared with NATs for all nine cancer types (all permute P-value < 0.001, Fig. 2a, b, Supplementary Fig. 1a and Methods). The most pronounced reduction of sex differences in DNAm was observed in BLCA, with the fewest sDMPs identified in the cancer state (Fig. 2a). These results demonstrate a marked attenuation of sex differences at DNAm levels in cancers, yielding a more epigenetically homogeneous pattern during cancer progression.
Fig. 2
Attenuation of sex differences in DNAm across nine cancers. a Overview of methods for identifying statistically significant differences in the number of sDMPs across nine cancers. The comparison of BLCA versus NAT is used here as an example. Left, the true difference in the number of sDMPs between BLCA and NAT samples. Right, a permuted null distribution (1000 times) is then used to determine the significance of the difference in the number of sDMPs between BLCA and NAT samples. b The true differences in the number of sDMPs between the paired cancers and NAT samples across all nine cancers. All pairs reaching a permutation P < 0.001. c The number of AMPs identified in each cancer. The number of female- (higher DNAm levels in females) and male-biased (higher DNAm levels in males) AMPs are indicated. d UpSet plot of AMPs’ overlap between pairs of cancers. Dark dots and lines indicate that the set participates in the intersection. The doughnut chart indicates the cancer sharing profile of AMPs. e, PC1 of DNAm levels of AMPs between cancer and NAT samples across all pairs. f The enrichment of X-linked AMPs across these five evolutionary strata, labeled as S1 (the most conserved region) to S5 (the most recent region), arranged in order from the distal end of the long arm to the short arm. *FDR < 0.05; **FDR < 0.01; ***FDR < 0.001. sDMPs sex-related differentially methylated positions, NAT normal adjacent tissues, AMPs attenuated methylated positions, PC1 principal component 1
To explore how sex-related DNAm changes were reduced in cancer, we employed a conservative filtering process to extract sDMPs exhibiting attenuation (Methods). To elucidate this procedure, we defined attenuated methylated positions (AMPs) as sDMPs that were significantly methylated in NATs but not in cancers. We identified 3,452 significant AMPs (Pbonf < 0.05), ranging from 426 to 1,040 AMPs per cancer (Fig. 2c and Supplementary Table 3). Regarding the direction of DNAm changes, 1,515 AMPs displayed higher methylation in males than in females and were classified as male-biased, while 1,970 AMPs showed higher methylation in females and were classified as female-biased. Overall, 63.12% (2,179/3,452) of these AMPs were cancer-specific (Fig. 2d and Supplementary Table 3). These AMPs exhibited greater DNAm changes in NATs than in cancers, representing the pattern of attenuation (Fig. 2e).
We found that 88.76% (3,064/3,452) of the AMPs were X-linked, a proportion that was not significantly different from the proportion of X-linked sDMPs in NATs (90.89%, 8028/8833; Fisher’s exact test, P = 0.23). Notably, 72% (442/614) of AMPs annotated genes were previously identified as X chromosome inactivation (XCI) genes, particularly the XCI inactive genes (Supplementary Fig. 1b, Supplementary Table 3 and 4). Positional enrichment on the X chromosome revealed that female-biased AMPs were strongly enriched in the X conserved region of the long arm in five out of nine cancers (HNSC, COAD, LIHC, BLCA, and LUSC), suggesting escape from XCI in cancers (false discovery rate (FDR)-corrected P < 0.05, Fig. 2f and Supplementary Table 4). Additionally, in contrast to male-biased AMPs, the detected female-biased AMPs predominantly resided in promoter and proximal enhancer regions, except for KIRC (FDR < 0.05, Supplementary Fig. 1c and Supplementary Table 4).
To investigate the biological processes contributing to this broad attenuation of sex differences in cancers, we performed functional enrichment analysis on the identified AMPs. Except for KIRC and THCA, we observed pervasive dysregulation of the negative regulation of transcription in the remaining seven types of cancers (Supplementary Fig. 1d and Supplementary Table 5). Notably, dysregulation of synapse-related pathways was identified in LUAD, LUSC, BLCA, COAD, and KIRP (Supplementary Fig. 1d and Supplementary Table 5). Emerging evidence indicates that crosstalk between the nervous system and cancer is a crucial regulator of the tumor microenvironment [54, 55]. These findings suggest that the sex differences in DNAm are reorganized in cancers, further contributing to the construction of the tumor microenvironment.
Sex-stratified differential DNAm was largely cancer-specificTo understand whether the alteration of DNAm in cancers involving sex-dependent regulation, a sex-stratified approach was applied to assess the differential methylation between cancers and NATs. We discovered a total of 36,693 female-related DMPs (fDMPs, Pbonf < 0.05 and permutation-based P < 0.05), with 177–9,025 fDMPs identified per cancer (Fig. 3a and Supplementary Table 6). Meanwhile, there were 88,811 male-related DMPs (mDMPs) identified (Pbonf < 0.05 and permutation-based P < 0.05), ranging from 931 to 44,030 mDMPs per cancer (Fig. 3a and Supplementary Table 6). Of these, essentially all (~ 99%) of the sex-stratified DMPs were located at autosomes (Fig. 3a). We quantified the replication rates for these sex-stratified DMPs in five external datasets for LIHC, LUAD, COAD, KIRC, and THCA and observed high replication rates (average π1 in females: 0.89; in males: 0.90; all spearman’s ρ > 0.70; Fig. 3b, Supplementary Fig. 2a and Supplementary Table 7).
Fig. 3
Sex-stratified cancer-related differential DNAm. a Discovery of sex-stratified DMPs across nine cancer types. Left, the number of sample sizes per sex and group (heatmap). Right, the number of identified fDMPs and mDMPs (Bonferroni-adjust P < 0.05 and permutation P < 0.05, histogram). Proportions of X-linked and autosomal f(m)DMPs (chr.) and of the overlap between fDMPs and mDMPs per cancer are indicated (stacked bar plots). b Replication rates (π1) for identified fDMPs and mDMPs in five types of cancers (LIHC, LUAD, COAD, KIRC, and THCA). c Cancer sharing profile of sex-stratified DMPs. d Absolute values of effect sizes for fDMPs and mDMPs. The dashed lines represent the median of effect sizes per sex. The FDR-corrected P-values according to paired Wilcoxon signed-rank tests are presented. fDMPs female-related DMPs, mDMPs male-related DMPs
Although the number of DMPs differed between males and females across cancers, an average of 23% of sex-stratified DMPs overlapped (Fig. 3a). The highest overlap was in COAD (43%), while BLCA had the lowest (3%; Fig. 3a). Furthermore, sex-stratified DMPs showed a pronounced cancer-specific pattern, with 77.41% of fDMPs and 67.07% of mDMPs being unique to individual cancers (Fig. 3c, Supplementary Fig. 2b).
The number of sex-stratified DMPs across cancers varies considerably, which may reflect sample sizes differences (Spearman’s ρ = 0.42, Fig. 3a). To ensure such sex bias were not driven by sample size differences, we performed 1000 rounds of down-sampling analyses and bootstrapping tests. Three types of cancers (COAD, LUAD, BLCA) exhibited significant sex differences in the number of DMPs based on the down-sampling analyses, with LUAD and COAD having more DMPs in females and BLCA having more DMPs in males (Supplementary Fig. 2c). Additionally, six other cancer types showed significant sex bias using the bootstrap-based approach (Supplementary Fig. 2d).
We next investigated whether the correlation and magnitude of DNAm changes differed by sex in cancers. The effect sizes were highly correlated between sexes across cancers, with ~ 99% of the sex-stratified DMPs showing consistent direction of DNAm changes (all Spearman’s ρ > 0.8, Supplementary Fig. 2e). Notably, the distribution of DNAm changes was significantly sex-biased across all nine cancers (paired Wilcoxon signed-rank test with FDR-corrected P < 0.05, Fig. 3d). The most prominent sex difference was observed in BLCA, with a significantly male-biased magnitude of DNAm changes (Fig. 3d). These results indicate largely consistent directional DNAm changes between sexes, but with varying magnitudes across cancers.
Sex-by-cancer interaction effects on DNAm were ubiquitous across cancersWe next evaluated the interaction effects between sex and cancer (cancer or NAT) across all nine cancers. There were 1,642 interaction DMPs identified, with 386 (23.51%) showing differential methylation in more than two different cancer types, suggesting cancer-dependent regulation (Fig. 4a, b and Supplementary Table 8). These interaction DMPs were largely X-linked (86.67%, Fig. 4a). Only a limited fraction of interaction DMPs overlapped with the identified sex-stratified DMPs, representing 15.11% for fDMPs and 30.49% for mDMPs on average (Fig. 4c). These suggest that these two analytical strategies complement each other and coordinately reflect the sex effects on DNAm in cancers.
Fig. 4
Sex-by-cancer interaction effects on DNAm. a Discovery of sex-by-groups interaction DMPs across nine cancer types. The proportions of X-linked and autosomal f(m)DMPs (chr.) per cancer are indicated (left, stacked bar plots). The number of interaction DMPs per cancer are shown (right, histogram). Legend colors are as in Fig. 3. b Cancer sharing profile of interaction DMPs. c Pie plots indicate the proportions of overlap between interaction DMPs and sex-stratified DMPs per cancer. d Cancer-related differential methylation effect size of females compared with males for the interaction DMPs per cancer. The slope (S) is calculated using principal components regression, with *FDR < 0.05 indicating that S is significantly different between sexes (see details in Methods). The dashed line represents equal effects between females and males when S = 1
We conducted functional enrichment analyses of the interaction DMPs across cancers, identifying significant enrichment in pathways associated with cell metabolism, cell cycle control, immune response, and tumor microenvironment interactions (Supplementary Table 8). Notably, synapse-related pathways were commonly dysregulated across most cancers, except THCA, underscoring the potential impact of sex effects on cancer-nervous system interactions.
To investigate the sex-specific effects of these interaction DMPs, we directly compared the DNAm changes of these loci in a sex-stratified manner. We found that the effect sizes of these interaction DMPs were significantly sex-biased in six out of nine cancers (Fig. 4d and Supplementary Fig. 2f). Specifically, THCA (Slope, S = 3.1, FDR < 0.001), COAD (S = 2.8, FDR < 0.001), HNSC (S = 2.3, FDR < 0.001), BLCA (S = 1.9, FDR = 0.039), KIRC (S = 1.8, FDR = 0.032), and LUAD (S = 1.5, FDR = 0.002) showed larger magnitude of changes in males than in females. These observations are consistent with the sex-stratified results, highlighting that sex effects on cancers may largely manifest through differences in the magnitude of multiple DNAm effects.
Amplification effects of DNAm account for sex differences in cancersOur observations highlight a pervasive sex difference in the magnitude of DNAm effects on cancers. However, involving dysregulation of multiple genes during cancer progression, how to quantify the extent of sex-dependent effects in cancers presents a challenge. Inspired by a recent study [35] that proposed sex differences in the magnitude of many genetic effects (‘‘amplification’’) as the primary mode of gene-by-sex interaction in complex human traits, we inferred that such a theory could be extended to DNAm levels in cancers. To evaluate the sex-dependent amplification effects in cancers, we examined the covariance relationships of DNAm effects between females and males using the multivariate adaptive shrinkage (mash) approach (Methods).
For these nine cancer types, we noticed that the DNAm effects exhibiting negative correlation (corr < 0) were more likely to be marginal effects, unable to survive the multiple testing correction penalty (Supplementary Fig. 3b). On average, 68% of DNAm effects showed perfect correlation (corr = 1) between sexes in cancers, with an additional 27% exhibiting partial correlation (0 < corr < 1) (Fig. 5a, Supplementary Fig. 3d and Supplementary Table 9). These suggest that most DNAm effects were perfectly or partially correlated. While negative correlation, representing opposite regulation of DNAm between sexes, may not be the primary driver of sex differences in cancers.
Fig. 5
Amplification effects on DNAm in cancers. a Proportions of weights on different types of correlation relationships between female and males for each cancer. Perfect correlation is indicated when corr = 1. Partial correlation is represented when 0 < corr < 1. Negative correlation is denoted when corr < 0. Uncorrelated condition is described when corr = 0. b Proportions of weights on different types of magnitude relationships between sexes for nine cancers. "female > male" indicates larger effect sizes in females than in males. "female < male" indicates larger effect sizes in males than in females. "female = male" represents equal effect sizes between sexes. c The difference between the fraction of male-larger effects and the fraction of female-larger effects classified these nine cancer types into female- and male-biased group. The male-biased group is located above the diagonal, while female-biased group is located below the diagonal. d Example of weights on covariance matrices for BLCA, which exhibited the greatest magnitude of DNAm changes in males
In terms of magnitude relationships, we found widespread sex-biased effect sizes across cancers. Specifically, on average, 34% of effects exhibited male-biased larger DNAm changes across the nine cancers, while 20% exhibited female-biased larger changes (Fig. 5b, Supplementary Fig. 3d and Supplementary Table 9). These findings align with our sex-stratified results, indicating that the cancer-related DNAm effects between sexes were largely correlated but with varying magnitudes.
We next classified these nine cancer types based on the difference between the fraction of male- and female-larger effects. The female-biased group included LUSC, LIHC, COAD, and KIRC, each exhibiting a relatively greater percentage of female-larger effects (Fig. 5c, Supplementary Fig. 3c and Supplementary Table 9). The male-biased group included BLCA, THCA, KIRP, LUAD, and HNSC, each showing a relatively higher percentage of male-larger effects (Fig. 5c, Supplementary Fig. 3c and Supplementary Table 9). Remarkably, BLCA displayed the greatest magnitude of DNAm changes in males. Of the 79% weights on matrices showing perfect correlation between sexes, 78% of the weights represented larger DNAm changes in males, with 46% exhibiting three times male-larger effects (Fig. 5d, Supplementary Fig. 3d and Supplementary Table 9). Together, these observations point to amplification effects of DNAm as the primary mode for sex differences in cancers.
DNAm signals exhibiting cancer-related sex amplification effectsWe then prioritized the hub DNAm signals representing the observed magnitude differences in cancer-related DNAm changes across sexes. We assessed the heterogeneity in effect sizes between sexes for each cancer and integrated these results with sex-stratified findings and posterior estimates assigned by the mash model (Supplementary Fig. 4; Methods). These analyses yielded 3,361 female-amplifiers, 11,837 male-amplifiers, and 25,911 sex-shared DNAm effects across nine types of cancers (Fig. 6a and Supplementary Table 10). No significant opposing effects between sexes were identified (Fig. 6a). These DNAm effects were largely (> 81%) cancer-specific (Fig. 6b).
Fig. 6
Prioritize DNAm signals exhibiting cancer-related sex amplification effects. a The number of cancer-related DMPs exhibiting the sex amplification effects (right, histogram). No significant opposite effects between sexes were identified. Proportions of X-linked and autosomal DMPs for sex-shared effects, female-amplifiers, and male-amplifiers (left). b Cancer sharing profile of sex-shared effects, female-amplifiers, and male-amplifiers. c Enrichment of genomic functional elements (CGI regions, genomic features, and cCRE regions) for sex-shared effects and sex-amplifiers. The signed odds ratios are labeled in colors (two-sided: hypermethylated and hypomethylated). Significant FDR-corrected P-values are indicated (*FDR < 0.05). d Sex-specific TFBS enrichment for sex-shared DMPs and sex-amplifiers. The TFs that were sex-specifically enriched in cancers are depicted (top). The TFBS enrichment profiles for each cancer are shown (bottom). The red words represent TFs with female-specific TFBSs enrichment, blue words represent TFs with male-specific TFBSs enrichment, and black words represent TFs with sex-biased TFBSs enrichment. The signed odds ratios are labeled in colors (two-sided: hypermethylated and hypomethylated). Significant FDR-corrected P-values are indicated (*FDR < 0.05). e The heatmaps illustrate the top three significantly estimated hazard ratios (HRs) for each cancer, which are associated with overall survival. *P < 0.05; **P < 0.01; ***P < 0.001. CGI CpG island, cCREs candidate cis-regulatory elements, TFBSs transcription factor binding sites
To determine the functional regulatory role of these sex-shared and sex-amplified effects in cancers, we performed enrichment analysis of genomic functional elements for these effects. We found that these effects were both significantly enriched in gene regulatory regions, particularly in gene body regions, untranslated exon regions (UTRs), and distal enhancer regions (Fig. 6c and Supplementary Table 11).
DNAm in enhancer regions is closely linked to regulating gene expression. We inferred that could sex-specifically affect the occupancy of transcription factors (TFs) at their binding sites (TFBSs). To test this, we examined the enrichment of 137 TFs for their binding sites in sex-shared and sex-amplified effects. We found that the enrichment for TFBSs were largely driven by hypomethylated DMPs (Fig. 6d and Supplementary Table 12). Among the tested TFs, 54.74% (75/137) were sex-specifically enriched in at least one cancer type, with TFBSs for 24 TFs implicated in two types (Fig. 6d). Such is the case of EGR2, which was specifically enriched in female-amplifiers of both LUAD and HNSC (Fig. 6d). A recent study highlighted the role of EGR2 in coordinating the alveolar macrophage functional program [56]. Our findings imply a female-specific role of EGR2 in LUAD under the influence of female-amplified aberrant DNAm. Moreover, 14 TFs displayed sex-biased roles in different cancers according to the enrichment profiles (Fig. 6d and Supplementary Table 12). For instance, CTCF was significantly enriched in male-amplifiers in COAD, whereas in female-amplifiers in LIHC. Additionally, to identify enriched conserved motifs within the X-linked sex-amplifiers in cancers, we used HOMER [48]. Our analysis revealed enrichment for three conserved TF motifs among the X-linked female-amplifiers and 66 motifs for the male-amplifiers (Supplementary Table 12). Notably, 17 TFs among the X-linked male-amplifiers exhibited motif enrichment across more than two cancer types, including AP-1, E2F1, and Sox7. Importantly, the enriched motifs for female- and male-amplifiers were distinct.
To understand the biological roles of sex-amplified DMPs in cancers, we performed functional enrichment analyses. For female-amplifiers, we observed dysregulation of immune-related pathways, including interleukin-18-mediated signaling in LIHC, type I interferon signaling in HNSC, and antigen presentation via MHC class II in BLCA (Supplementary Fig. 5a and Supplementary Table 13). In male-amplifiers, pathways related to cell proliferation, migration, and TGF-β signaling were affected, such as negative regulation of endothelial cell proliferation in LUSC, regulation of cell migration in KIRC and THCA, and response to TGF-β in BLCA (Supplementary Fig. 5b and Supplementary Table 14).
Given the pervasive enrichment of synapse-related pathways among AMPs, we examined sex-amplified genes within synaptic biological functions using the SynGO database [57]. We found that 244 female-amplified genes and 540 male-amplified genes were annotated with synaptic functions. Although synaptic process enrichment is broadly consistent between sexes, differences exist in the specific biological terms and genes mapped within the SynGO hierarchy (Supplementary Fig. 5c and Supplementary Table 15). For example, male-amplified genes such as DLG3, PORCN, and PPFIA1 are involved in regulating postsynaptic membrane neurotransmitter receptor levels across multiple cancer types. In contrast, a distinct set of female-amplified genes, such as NRXN3 and EFNB2, are linked to the same pathway in comparable cancer types. Additionally, male-amplified genes EXOC2, ATG16L1, and NETO1 are associated with axonal transport in BLCA, HNSC, COAD, and KIRP, indicating potential male-specific pathological mechanisms. The diversity of synaptic functions among sex-amplified genes implies that various functional interactions between sex and cancer risk converge at the synapse.
To demonstrate the functional and clinical relevance of these DNAm divergences, we assessed the association of sex-amplifiers with overall patient survival and recurrence following initial treatment. Using Lasso regression for variable selection followed by multivariate Cox analysis, we identified 24 female-amplifiers and 29 male-amplifiers that were sex-specifically associated with overall survival (Fig. 6e and Supplementary Table 15). For example, the female amplifier for BLCA, cg19395441, was significantly associated with overall survival in females (HR = 1.48, 95% CI 1.21–1.82, P < 0.001) but not in males (HR = 0.96, 95% CI 0.86–1.07, P = 0.48). Additionally, we identified 15 female-amplifiers and 13 male-amplifiers associated with recurrence (Supplementary Table 15). These findings highlight the potential of identified sex-amplifiers as the sex-specific prognostic biomarkers.
Integration of DNA methylation and gene expression highlights sex-dependent regulation and biological processesThe primary function of DNAm is to regulate gene expression. To gain insight into its potential regulatory role, we characterized the correlations between methylome and transcriptome (eQTMs) in each sex per cancer, using matched samples. A total of 38,969 eQTMs were identified in females, ranging from 1,135 in KIRP to 17,685 in LUAD (Spearman’s Pbonf < 0.05, Fig. 7a and Supplementary Table 16). Meanwhile, 77,930 eQTMs were observed in males, ranging from 2,770 in THCA to 28,605 in BLCA (Spearman’s Pbonf < 0.05, Fig. 7a and Supplementary Table 16). On average, 24.17% of eQTMs were shared between sexes with consistent directions of correlation (Fig. 7a), and none of the eQTMs exhibited opposite correlations between sexes. Among these eQTMs, 71.74% (27,958/38,969) of female-related eQTMs and 56.19% (43,788/77,930) of male-related eQTMs were significantly correlated in only one type of cancer, indicating cancer-dependent regulation (Fig. 7b). The prevalence of sex-specific eQTMs across cancers highlights extensive sex-dependent regulatory mechanisms.
Fig. 7
Characterizing the correlations between DNAm and gene expression. a Discovery of eQTMs pairs per sex and per cancer. Left, the number of sample sizes per sex and per cancer (heatmap). Right, the number of eQTMs (Bonferroni-adjust P < 0.05, histogram). The sex-overlapped eQTMs, female-specific eQTMs, and male-specific eQTMs are indicated. b Cancer sharing profile of eQTMs per sex. c Discovery of the detected sex-amplifiers with significantly sex-stratified differential gene expression using eQTM pairs. The directions of correlation are depicted. d Examples of female-amplifiers that were significantly correlated with gene expression of target genes, exhibiting a consistently greater magnitude of gene expression changes in females compared to males. e Examples of male-amplifiers that were significantly correlated with gene expression of target genes, exhibiting a consistently greater magnitude of gene expression changes in males compared to females. *Pbonf < 0.05; **Pbonf < 0.01; ***Pbonf < 0.001. eQTMs methylome and transcriptome correlations
To investigate the functional mechanisms underlying these eQTMs, we performed enrichment analysis of candidate cis-regulatory elements (cCREs) for eCpGs in negatively and positively correlated eQTMs per sex, respectively. A similar enrichment pattern was observed in female- and male-related eQTMs. In both sexes, positively correlated eQTMs were more frequently enriched in distal enhancer regions and insulators, whereas negatively correlated eQTMs were predominantly enriched in promoters and proximal enhancer regions (Supplementary Fig. 6a and Supplementary Table 17).
We next prioritized eQTM pairs associated with sex-amplified DMPs. For female-amplifiers, 48 eQTMs were extracted across five types of cancers (number of eQTMs in descending order; LUAD: 27, LIHC: 8, COAD: 7, KIRC: 4, and THCA: 2, Fig. 7c and Supplementary Table 18). These eQTMs comprised 44 eCpGs and 42 eGenes, with a majority (68%, 33/48) exhibiting negative correlations (Fig. 7c). Consistent with DNAm patterns, 32 eGenes demonstrated greater magnitudes of expression changes in females compared to males (Supplementary Table 18). Notably, in LIHC, three female-amplifier CpGs (cg01714932, cg08729600, and cg10479063) were significantly negatively correlated with PZP gene expression (Spearman’s Pbonf < 0.05), with larger expression changes in females (Fig. 7d and Supplementary Table 18). Additional notable eQTM pairs for female-amplifiers included cg17306740-ZBP1 in LUAD, cg01765174-TRIM14 in COAD, and cg25748357-GRB10 in KIRC (Supplementary Fig. 6b-d and Supplementary Table 18).
Regarding the male-amplifiers, 380 eQTMs were extracted for eight cancer types linked to 356 eCpGs and 272 eGenes (number of eQTMs in descending order; KIRP: 183, HNSC: 68, KIRC: 40, LUSC: 34, LIHC: 27; COAD: 15, LUAD: 12, THCA: 4, Fig. 7c and Supplementary Table 18). Of these, 277 eQTMs exhibited negative correlations (Fig. 7c), with 285 showing consistently greater magnitudes of DNAm and gene expression changes in males. For instance, the male-amplifier cg12560128 was negatively correlated with OAS2 and OAS3 in COAD (Fig. 7e and Supplementary Table 18), both known immune biomarkers linked to the tumor microenvironment [58, 59]. Additionally, HRH1, which correlated negatively with cg17660833 in KIRC, has been implicated in T cell dysfunction and is frequently upregulated in the tumor microenvironment [60]. (Fig. 7e and Supplementary Table 18). Additional examples include cg06490988-GATA2 in KIRP, cg19193956-TWIST1 in HNSC, cg13784235-NR2E1 in LUSC and cg15652212/cg22531018/cg21852589-TMEM139 in LIHC (Supplementary Fig. 6e-j and Supplementary Table 18). These results suggest that sex-amplified DNAm changes can influence downstream gene expression, contributing to consistent sex-biased differential expression in cancers.
To elucidate the biological roles of genes regulated by sex-amplifiers through eQTMs, we performed functional enrichment analysis. Our findings indicate that genes regulated by male-amplifiers are associated with cancer development-related pathways, including vasculature development, epithelial cell differentiation, and myeloid cell differentiation (Supplementary Fig. 7 and Supplementary Table 19). In contrast, genes regulated by female-amplifiers are linked to specific pathways, such as sprouting angiogenesis in COAD and cell communication in THCA (Supplementary Fig. 7 and Supplementary Table 19).
We further evaluated the therapeutic potential of genes regulated by sex-amplifiers through eQTMs. Using druggable targets annotated in the CIViC database [53], we found that several genes were previous identified as drug targets (Supplementary Table 18). For instance, ECSCR in LUAD and PTK2B (also known as PYK2) in COAD were regulated by female-amplifiers, while GATA2 in KIRP, ERBB3 in HNSC, and POU5F1 in KIRC were regulated by male-amplifiers. These observations indicate that these previously identified druggable targets may have a sex-specific role in cancers.
留言 (0)