ASIC1A was upregulated in NSCs following SCI
To address whether ASIC1A was upregulated following SCI, we detected the expression of ASIC1A within the lesion sites at different time points post-injury by western blot. The expression level of ASIC1A was increased at 1 day post-injury (dpi), reached its peak at 3 dpi, and then gradually decreased (Fig. 1A, B). To further investigate which type of nerve cells expressed ASIC1A, we co-labeled ASIC1A with different nerve cells by immunostaining in SCI rats. Given that the highest expression of ASIC1A was observed at 3 dpi, we focused on this time point for immunostaining. Based on the characteristics of cell biology following SCI, the injured spinal cord was divided into three regions: the lesion core (LC), the edge of the lesion core (ELC), and the surrounding neural tissue (SNT) (Fig. 1C). Compared with the SNT, which was remote from the LC, the ASIC1A expression level was strongly increased in the LC and ELC regions. In these two regions, ASIC1A was found to be colocalized with NSCs, NG2-positive oligodendrocyte precursor cells (OPCs), and Tuj-1-positive neurons (Fig. 1D–F). In the LC, 10% of ASIC1A and Nestin double-positive NSCs (accounting for 72.8% of total NSCs, Fig. 1D, I, J) and 3.2% of ASIC1A and NG2 double-positive OPCs (accounting for 48.9% of total OPCs) were found in this region (Fig. 1E, I, K). Although 56% of TUJ1-positive neurons (Fig. 1L) were found to express ASIC1A, due to the initial damage-induced loss of neurons, only a few neurons were found in this region, resulting in an extremely low percentage of TUJ1- and ASIC1A-double-positive cells (Fig. 1F, I). In the ELC, ASIC1A was also found to be colocalized mainly with NSCs, OPCs and TUJ-1-positive neurons. The double-positive percentage was 13.4% in NSCs (accounting for 46.2% of total NSCs, Fig. 1D, I, J) and 8% in NG2+ OPCs (accounting for 38.7% of total OPCs, Fig. 1E, I, K). With an increased number of TUJ1-positive neurons in the ELC, the percentage of TUJ 1- and ASIC1A-double-positive cells rose to 11.5% (Fig. 1F, I), accounting for 38.8% of the total neurons (Fig. 1L). In the SNT, where ASIC1A expression was considerably reduced, there was a corresponding decrease in the percentage of ASIC1A-positive NSCs, OPCs and neurons (Fig. 1D–F, I–L). In contrast, in these three regions, few GFAP-positive astrocytes and CNPASE-positive oligodendrocytes co-expressed ASIC1A (Fig. 1G–I).
Although Nestin was highly expressed in NSCs, it was not a specific marker to identify NSCs. It has been reported to be expressed in reactive astrocytes [35]. To confirm whether these Nestin-positive cells were from reactive astrocytes, we co-labeled ASIC1A with Nestin and GFAP by immunostaining in ELC at 3 dpi. The results showed that among the Nestin- and ASIC1A-double positive cells, only 20% cells were co-labeled with GFAP, indicating that most of the Nestin-positive cells were not from reactive astrocytes (Fig. 1M, N). Next, Sox 2, which is essential for maintaining pluripotency in stem cells and highly expressed in NSCs [36, 37], was used to co-label with ASIC1A and Nestin. It showed that nearly 75% ASIC1A- and Nestin-double positive were co-labeled with Sox 2 (Fig. 1O, P). These results indicated that ASIC1A was markedly upregulated in NSCs following SCI.
ASIC1A activation prevents NSCs from differentiating into oligodendrocytes
To confirm whether ASIC1A expression in NSCs was activated by the extracellular acidic microenvironment within lesion sites following SCI, we first detected the tissue pH alteration in the lesion site (1 cm centered on the epicenter of the compressed lesion) and adjacent segment (1 cm from the cranial edge of lesion site) following SCI by a micro–pH meter. Consistent with previous studies [38, 39], we found a lower tissue pH value in both lesion site and adjacent segment than in sham group and persisted up to 7d following injury (Fig. 2A, B). The lowest pH (6.03 ± 0.06) value was noted in lesion site at 1 dpi (Fig. 2A). Due to the lowest pH value was approaching 6.0, pH 6.0 acidic medium was selected to culture with NSCs to mimic acidic microenvironment. Western blot results showed that the ASIC1A expression level in NSCs peaked at 24 h following exposure to pH 6.0 acidic medium (Fig. 2C, D). Double immunostaining of NESTIN and ASIC1A revealed a significant increase in the percentage of ASIC1A- and NESTIN-positive cells in NSCs treated with pH 6.0 acidic medium for 24 h compared with controls (Fig. 2E, F). To further investigate whether the degree of pH value affect the expression of ASIC1A in NSCs, NSCs were cultured with pH 6.5 acidic medium for 24 h and immunostaining was performed to detect the expression of ASIC1A. As expect, the expression of AISC1A was increased by the treatment of pH 6.5 acidic medium, but this effect was weaker than in pH 6.0 acidic medium-treated NSCs (Fig. 2E, F).
Since NSC differentiation is crucial for myelin sheath regeneration and neurological functional recovery, we examined whether acidic medium affects NSC differentiation. NSCs were digested into single cells and treated with normal culture medium or pH 6.0 acidic medium or pH 6.5 acidic medium for 5 days. Treatment of NSCs with pH 6.0 or pH 6.5 acidic medium resulted in a reduction in the percentage of CNPASE-positive oligodendrocytes and an increase in the proportion of GFAP-positive astrocytes compared with controls (Fig. 2G, H). In contrast, pH 6.0 acidic medium had a stronger effect on promoting the differentiation of NSCs into astrocytes, compared with pH 6.5 acidic medium (Fig. 2G, H). Moreover, to identify whether this acidic medium-induced effect was related to ASIC1A, the addition of the nonspecific ASIC antagonist amiloride and the ASIC1A antagonist psalmotoxin (PcTx) were used to inhibit ASICs and ASIC1A, respectively. As expected, the effect induced by acidic medium on NSC differentiation was abolished by amiloride or PcTx treatment (Fig. 2 G, H). Together, these data indicated that the acidic environment promoted NSC differentiation into astrocytes by upregulating the ASIC1A signaling pathway following SCI.
As extracellular acidification could directly induce apoptosis of cells, we next detected the apoptotic rate of NSCs following exposure to pH 6.0 acidic medium for 24 h by TUNEL staining. It revealed that the treatment of acidic medium significantly increased the apoptotic rate of NSCs compared with controls (Fig. 2I, J). Therefore, there is a possibility that the extracellular acidification might cause distinct apoptotic responses in oligodendrocytes and astrocytes, which resulted into a greater reduction in the proportion of oligodendrocytes during the process of NSC differentiation. To study this possibility, we cultured primary astrocytes and oligodendrocytes and treated these cells with pH 6.0 acidic medium for 24 h respectively. The TUNEL staining results showed that although compared with control groups, the apoptotic rates in oligodendrocytes and astrocytes were marked increased by the treatment of acidic medium, no difference were found between these 2 groups (Fig. 2I, J). It indicated that when facing extracellular acidification, the apoptosis of oligodendrocytes resembled that of astrocytes. These data confirmed that the extracellular acidification induced the reduction in the proportion of oligodendrocytes during the process of NSC differentiation mainly through preventing NSCs differentiation into oligodendrocytes rather than increasing the apoptosis of oligodendrocytes.
ASIC1A upregulated PTGS2 in acid-treated NSCs and SCI rats
To elucidate the molecular mechanism through which extracellular acidic environment influences the differentiation of NSCs, we conducted RNA-sequence analysis on NSCs treated with pH 6.0 acidic medium. Compared with controls, acidic medium treatment led to significant changes in gene expression, with 42 genes being upregulated and 192 genes being downregulated (Fig. 3A). Among the top ten upregulated and top ten downregulated genes, prostaglandin-endoperoxide synthase 2 (Ptgs2) stood out (Fig. 3B), and it is known for its crucial roles in post-trauma inflammation and demyelinating neurological diseases [40,41,42,43,44]. Moreover, KEGG pathway enrichment analysis showed that ptgs2 were related with the immune-relevant pathways (TNF-signaling pathway and IL-17 signaling pathway, Fig. 3C). By qPCR, we verified that ptgs2 gene expression was upregulated in NSCs following 24 h of exposure to acidic medium (Fig. 3D). Additionally, acidic medium treatment significantly increased the percentage of PTGS2 and NESTIN double-positive cells suggesting that an acidic environment could upregulate PTGS2 expression in NSCs (Fig. 3E, F). As expected, compared with pH 6.5-acidic medium treated NSCs, pH 6.0-acidic medium revealed a stronger effect on increasing the percentage of PTGS2 and NESTIN double-positive cells (Fig. 3E, F). To identify whether the upregulation of PTGS2 was related to ASIC1A, NSCs were treated with amiloride or PcTx to inhibit the activation of ASIC1A in an acidic environment, and the percentage of PTGS2 and NESTIN double-positive cells was strongly decreased following 24 h of culture compared with acidic medium-treated NSCs (Fig. 3E, F). These data suggested that the upregulation of PTGS2 in NSCs was related to extracellular acidic environment-induced ASIC1A activation. To further investigate whether the acidic environment-induced regulation of NSC differentiation was associated with PTGS2, we added NS398 (PTGS2 inhibitor) to NSCs in an acidic environment. This led to a significant weakening of the acidification-related effect on NSC differentiation, resulting in an increase in the proportion of oligodendrocytes and a reduction in the proportion of astrocytes (Fig. 3G, H). This result indicated that PTGS2 plays a key role in the extracellular acidic environment-induced regulation of NSC differentiation.
To further investigate the relationship between PTGS2 and ASIC1A in SCI rats, we assessed PTGS2 expression in SCI rats at 3 and 7 dpi by immunostaining. The results revealed that PTGS2 expression was significantly increased in the LC and ELC compared to the SNT. Moreover, many of these PTGS2-positive cells were also co-expressed with NESTIN-positive NSCs (Fig. 3I–K), indicating that injury to the spinal cord upregulated PTGS2 expression in NSCs. Next, to confirm whether the upregulation of PTGS2 was related to the activation of ASIC1A, we treated SCI rats with different concentration of amiloride or PcTx by a 3 day continuous intrathecal injection. It revealed that the higher dose of amiloride or PcTx appeared to provide stronger effects in inhibiting the expression of PTGS2 in NSCs at 3 dpi (Fig. 3I, J). No differences were found between the groups treated with amiloride at 100 μM and 200 μM or between the groups treated with PcTx at 10 nM and 20 nM (Fig. 3I, J). Therefore, 100 μM amiloride and 20 nM PcTx were selected for further injection. Similarly, this intervention significantly reduced the proportion of PTGS2-positive NSCs at 7 dpi (Fig. 3I, K). These in vivo results align with the in vitro findings, suggesting that inhibiting the activation of ASIC1A affects the upregulation of PTGS2 in NSCs, strongly indicating that the extracellular acidic environment upregulates PTGS2 expression via ASIC1A.
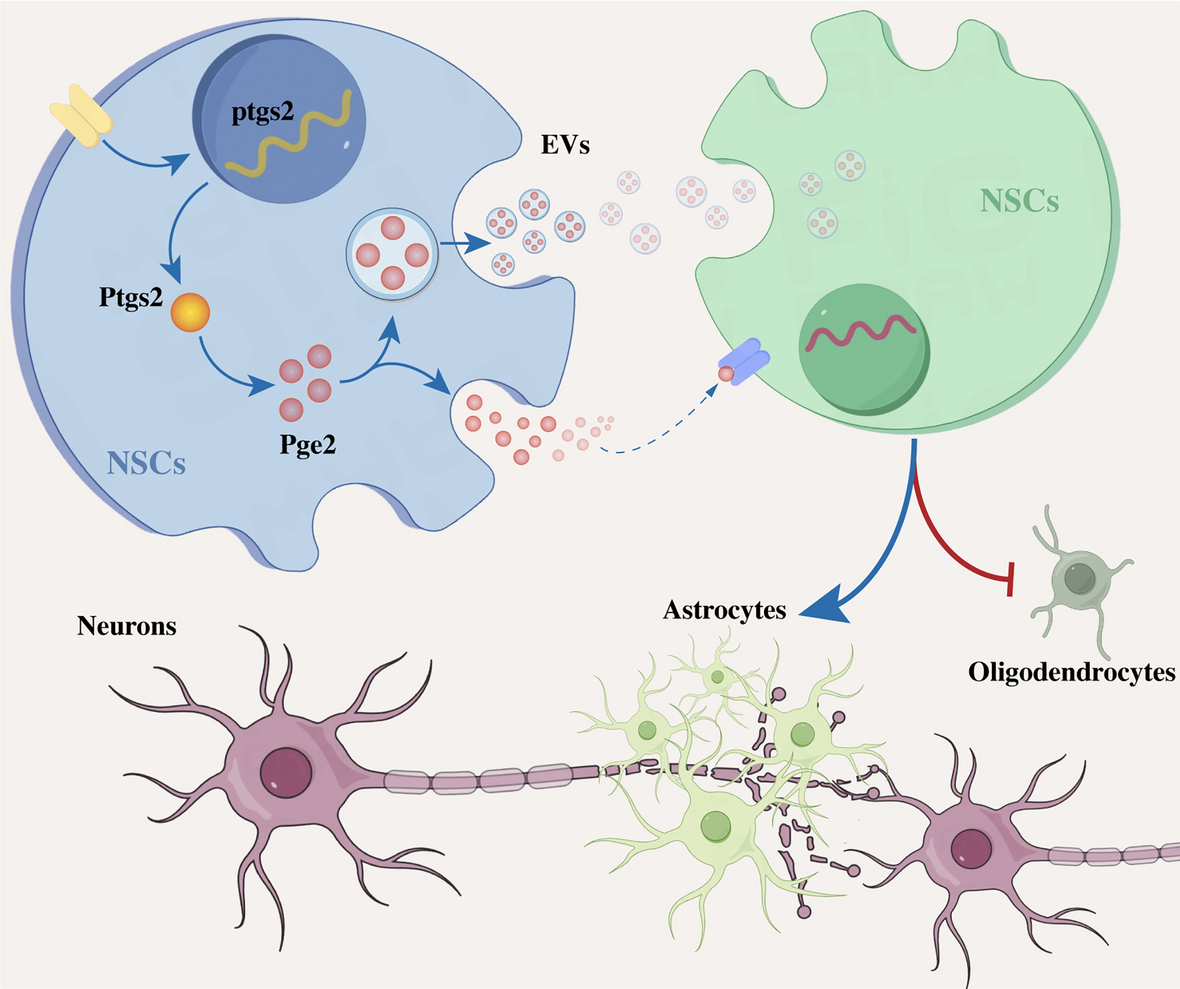
The acidic environment regulated NSC differentiation by enhancing transcellular NSC-to-NSC delivery of PGE2
PTGS2 plays a critical role in converting arachidonic acid into prostaglandin H2, which serves as the precursor for various prostaglandins, including prostaglandin E2 (PGE2) [45, 46]. PGE2 is a proinflammatory factor that can be secreted by both nerve and immune cells [47,48,49,50]. Additionally, PGE2 has been found to be enriched within extracellular vesicles (EVs) [51, 52], which are membrane-bound particles ranging from 30 to 150 nm and are released by a wide range of cell types [53, 54]. Our recent study has shown that endogenous NSCs can engage in cell-to-cell communication by releasing EVs [33]. Therefore, we hypothesized that in an acidic environment, NSCs release PGE2 within EVs and that these PGE2-enriched EVs regulate the differentiation of NSCs through intercellular communication. To confirm this hypothesis, we first collected EVs from NSCs and confirmed their identity by transmission electron microscopy (TEM), dynamic light scattering and western blot analysis (Fig. 4A–C). Subsequently, the concentration of PGE2 within EVs derived from NSCs treated with either normal medium (nm-NSC-EVs, control groups) or acidic medium (am-NSC-EVs, pH 6.0 groups) was determined using ELISA. Compared with untreated NSCs (controls), exposure to acidic medium at pH 6.0 significantly increased the concentration of PGE2 within EVs (Fig. 4D).
Since Pge2 facilitates signals by binding to specific cell surface prostaglandin E receptors (Ep1-Ep4) [55,56,57] and EP2 has been reported to be widely expressed in the central nervous system (CNS) [46, 58, 59], we then detected whether extracellular acidification could activate EP2 in NSCs. Following 24 h of treatment with acidic medium, immunostaining revealed that EP2 was colocalized with Nestin-positive NSCs, and the percentage of NESTIN- and EP2-double-positive cells markedly increased compared with controls (Fig. 4 E, F). Next, to confirm whether the upregulation of PGE2 and its related receptor EP2 were induced by activation of ASIC1A, we added PcTx to NSCs in an acidic environment. We then detected the concentration of PGE2 within NSC-EVs by ELISA and calculated the proportion of NESTIN- and EP2- double-positive cells by immunostaining. As expected, treatment with PcTx strongly reduced the concentration of PGE2 within EVs (Fig. 4D) and the percentage of double-positive cells compared to those exposed to acidic treatment at pH 6.0 (Fig. 4E, F). Moreover, in vivo, PcTx treatment reduced the expression of EP2 in NESTIN-positive NSCs in the LC and ELC at 3 and 7 dpi (Fig. 4G–I). These data indicated that acidic-dependent ASIC1A enhances the expression of PGE2/EP2.
To further investigate whether this ASIC1A-induced upregulation of PGE2 in NSC-EVs was associated with the regulation of NSC differentiation, we first treated NSCs with AH-6809 (a prostaglandin E2 receptor antagonist) in an acidic environment. This led to an increase in the percentage of CNPASE-positive cells compared with NSCs cultured with pH 6.0 acidic medium (Fig. 4J, K). To identify whether these acidic environment-induced alterations in NSC differentiation were related to NSC-EVs, we labeled NSC-EVs with Pkh-26 and treated NSCs and SCI rats with these Pkh-26-tracked NSC-EVs. Following a 24 h coculture with NSCs, it was discovered that labeled EVs (red) were present in the cytoplasm near the nucleus of NSCs (Fig. 4L). In vivo, following the intrathecal injection of Pkh-26-tracked NSC-EVs into SCI rats, immunostaining showed that these Pkh-26-tracked NSC-EVs were contained within the cytoplasm of these Nestin-positive cells (Fig. 4M).
Next, we collected EVs from normal medium-treated NSCs (nm-NSC-EVs) and acidic medium-treated NSCs (am-NSC-EVs), respectively, and treated NSCs with PGE2, nm-NSC-EVs and am-NSC-EVs for 5 days, and assessed the proportion of CNPASE-positive oligodendrocytes by immunostaining. Compared with the controls, the addition of nm-NSC-EVs significantly increased the proportion of CNPASE-positive cells (Fig. 4N, O). In contrast, the addition of PGE2 or am-NSC-EVs reduced the proportion of CNPASE-positive cells (Fig. 4N, O). Subsequently, we added PGE2 to NSCs in the presence of nm-NSC-EVs and AH-6809 to NSCs in the presence of am-NSC-EVs and showed that the addition of Pge2 abolished the nm-NSC-EV-induced effects on the differentiation of NSCs, resulting in a reduction in the proportion of CNPASE-positive cells (Fig. 4N, O). As expected, the addition of AH-6809 blocked the am-NSC-EV-induced effects and led to an increase in the percentage of CNPASE-positive cells (Fig. 4N, O). All these data indicate that the ASIC1A-induced upregulation of PGE2 within NSC-EVs plays a key role in preventing NSC differentiation toward oligodendrocytes.
Ptgs2 or Asic1a deficiency in NSCs countered acidic environment-induced effects on NSC differentiation by reducing the transcellular delivery of PGE2
To further confirm whether the am-NSC-EV-induced effect on mediating NSC differentiation was associated with PTGS2, siRNA-Ptgs2 (si-Ptgs2) was used to knock down Ptgs2 expression in NSCs (Fig. 5A). Compared with am-NSC-EVs, the concentration of PGE2 within the EVs derived from si-Ptgs2-transfected NSCs exposed to acidic medium (am-NSCsi-Ptgs2-EVs) was markedly reduced (Fig. 5B). As expected, am-NSCsi-Ptgs2-EVs lost their ability to regulate NSC differentiation, leading to an increase in the proportion of CNPASE-positive cells compared with NSCs treated with am-NSC-CM (Fig. 5C, D).
Since the upregulation of PTGS2/PGE2 was induced by the activation of ASIC1A, we used Asic1a-siRNA to knock down Asic1a expression in NSCs (Fig. 5E). In comparison with am-NSC-EVs, treatment with acidic medium did not enhance the concentration of PGE2 within EVs derived from si-Asic1a-transfected NSCs (Fig. 5F). Similarly, these EVs derived from am-treated NSCsi-Asic1a (am-NSCsi-Asic1a-EVs) resulted in an increase in the percentage of CNPASE-positive cells compared with am-NSC-EV-treated NSCs (Fig. 5G, H). All these findings indicated that the Asic1a-related delivery of PGE2 played a crucial role in preventing NSCs from differentiating into oligodendrocytes.
PcTx promoted myelin sheath regeneration and axonal remyelination in lesion sites
To evaluate the effects of PcTx on SCI rats, we first detected the regeneration of the myelin sheath at week 2 post-injury. Immunostaining showed that in SCI rats, a cavity formed in the center of the lesion site, surrounded by a GFAP-positive astrocytic boundary. Few CNPASE-positive cells were found within this astrocytic boundary and cavity. Rats injected with PcTx exhibited a significant increase in the percentage of CNPASE-positive areas (Fig. 6A, B), indicating enhanced myelin sheath regeneration.
As the corticospinal tract is of the utmost importance for the recovery of impaired motor function following SCI [60, 61], we detected axonal remyelination in the lateral corticospinal tract at week 4 post-injury by immunostaining with a mature oligodendrocyte marker, MBP. In SCI rats, regenerated TUJ1-positive neuronal fibers were observed around the cavity. However, compared with Sham rats, in which TUJ1-positive fibers were ensheathed with MBP-positive myelin sheets, the intensity of MBP-positive myelin within axons was significantly reduced in SCI rats (Fig. 6C, D), indicating demyelination in the axons. In contrast, injection of PcTx markedly increased the intensity of MBP-positive myelin sheets within the TUJ-positive areas (Fig. 6C, D), indicating that the injection of PcTx promoted axonal remyelination in lesion sites following SCI. The neurological results were consistent with the histological findings. The rats that received PcTx injection exhibited a better outcome in both BBB scores and angle of inclined plane test (Fig. 6E, F), indicating improved neurological functional recovery following SCI. All these data suggested that the PcTx treatment promoted myelin sheath regeneration and axonal remyelination, resulting in an improvement of the neurological functional recovery following SCI.
Asic1a-KO mice fail to upregulate PTGS2 expression, resulting in an improvement in axonal remyelination following SCI
To further study the role of ASIC1A in SCI and its relationship with PTGS2, Asic1a knockout (Asic1a-KO) mice were used (Fig. 7A). Following SCI in Asic1a-KO mice, PTGS2 expression in NSCs remained at a low level compared to that in wild-type (WT) mice (Fig. 7B, C). Next, NSCs were obtained from WT and Asic1a-KO mice and treated with acidic medium at pH 6.0, and their EVs were collected. Immunostaining showed that NSCs obtained from Asic1a-KO mice had a complete loss of Asic1a expression (Fig. 7D) and a lower percentage of PTGS2-positive cells than NSCs collected from WT mice (Fig. 7E, F). The ELISA results showed that the concentration of PGE2 was markedly lower in the EVs derived from Asic1a-KO NSCs (Fig. 7G). All these findings indicated that ASIC1A was needed for the upregulation of PTGS2 in response to the acidic environment.
To investigate the role of ASIC1A in NSC differentiation and axonal remyelination, NSCs obtained from WT mice and Asic1a-KO mice received acidic medium for 5 days. Immunostaining showed that Asic1a-KO NSCs had a higher percentage of CNPASE-positive NSCs than WT NSCs (Fig. 7H, I). Moreover, this Asic1a-KO-induced increase was abolished by the addition of PGE2, resulting in a decrease in the percentage of CNPASE-positive cells (Fig. 7H, I). These findings strongly indicated that PTGS2 played a key role in ASIC1A-induced mediation of NSC differentiation. At the edge of the lesion core, the total TUJ1 and MBP double-positive areas were significantly higher in Asic1a-KO mice than in WT mice at 4 weeks post-injury (Fig. 7J, K). Consistent with the histology results, Asic1a-KO mice exhibited better outcomes in both BBB scores and the inclined plane test (Fig. 7J, K). This highlighted that Asic1a deficiency enhanced axonal remyelination, leading to neurological recovery.




留言 (0)