
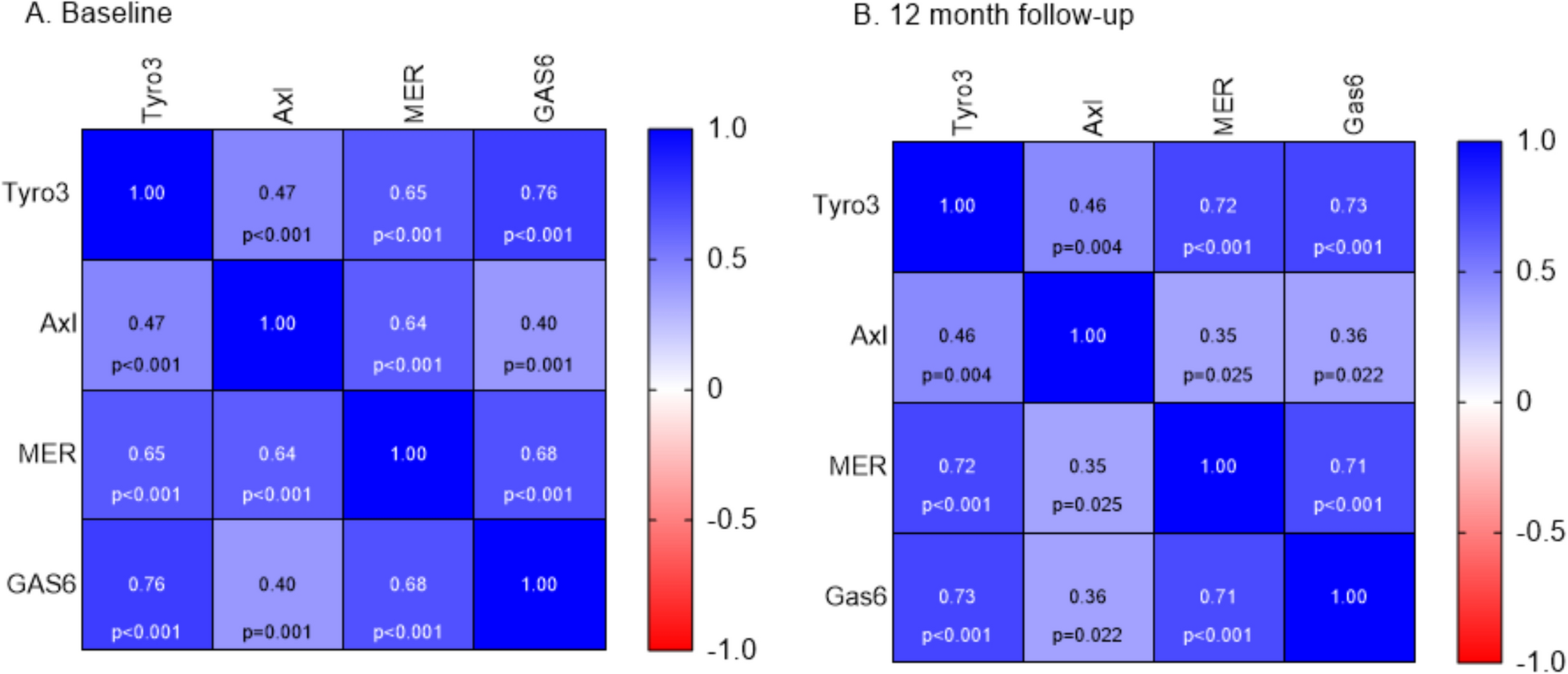
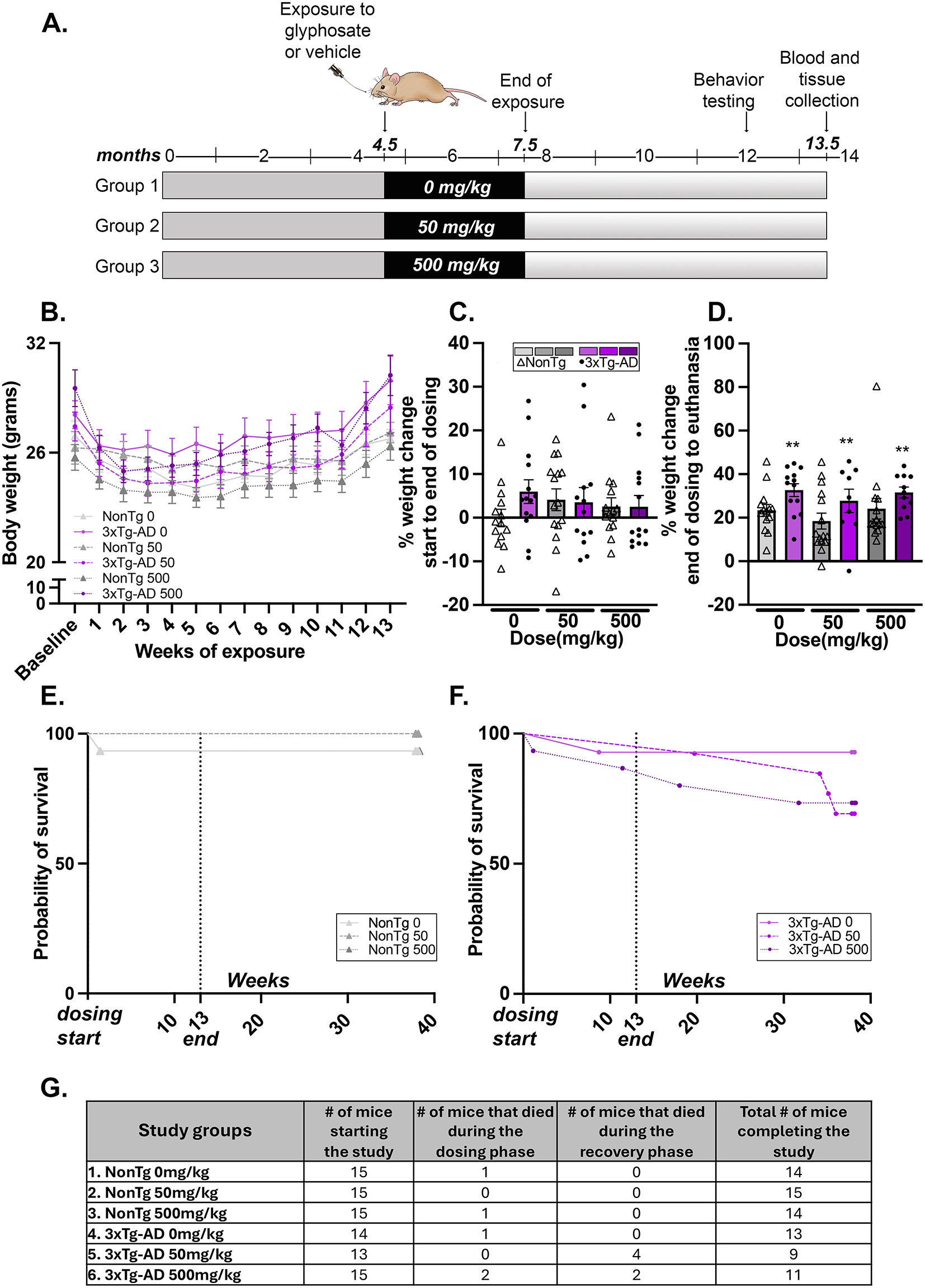
記住我

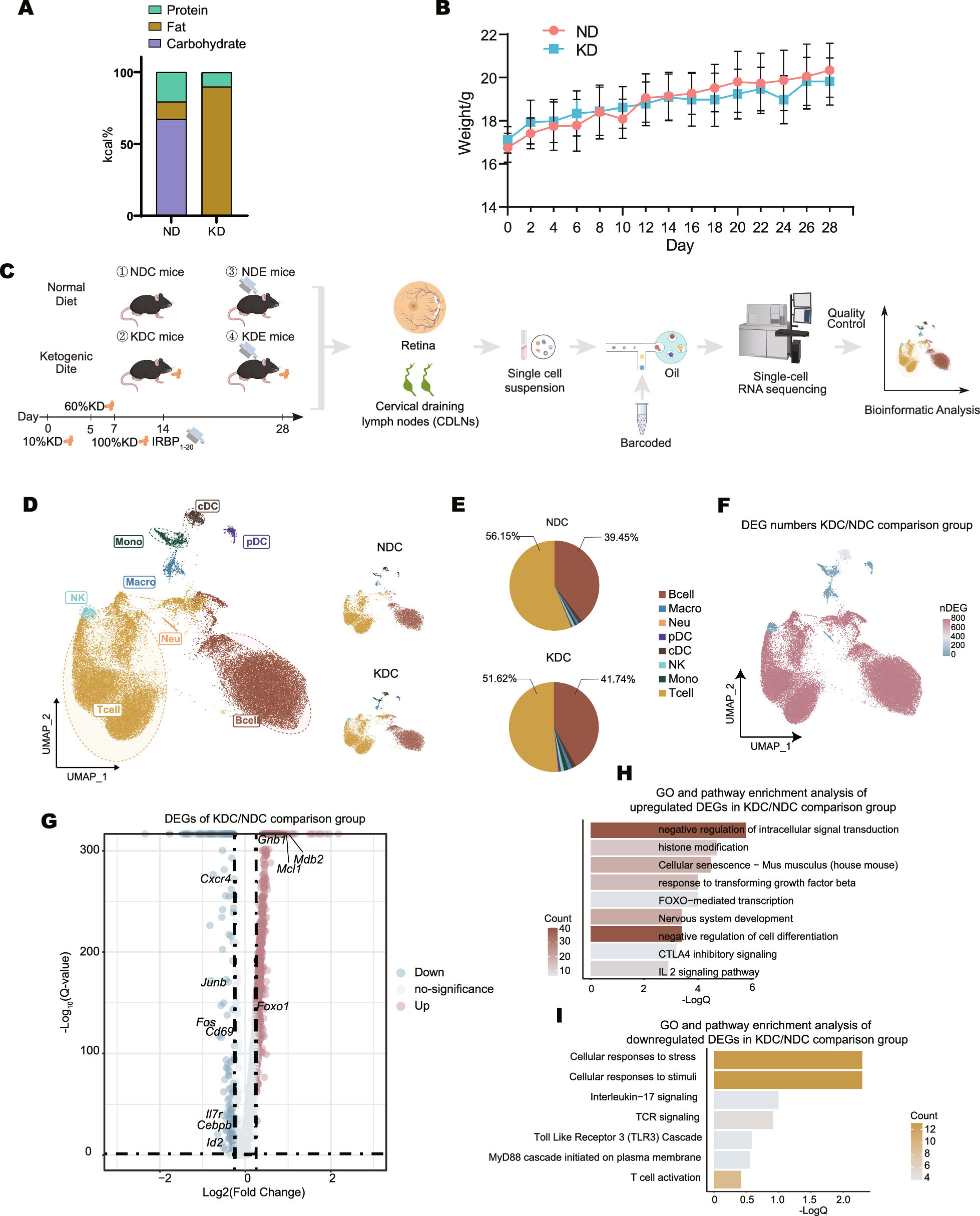
To evaluate the impact of ketogenic diet (KD), mice were evenly matched and then randomly allocated into two dietary cohorts: the normal diet (ND) and KD groups (Fig. 1a). Throughout the observation period, no significant difference in body weight (Fig. 1b) was observed between ND and KD group, indicating a comparable baseline nutritional status in both sets.
Fig. 1
Ketogenic diet induced complicated and extensive changes in the immune profile of CDLNs. a Daily diet composition during the normal diet (ND) and ketogenic diet (KD) stages respectively, represented in percent calories. b Changes in body weight of mice fed with ND or KD throughout the study protocol. c Schematic of the experimental design for single-cell RNA sequencing (scRNA-seq). CDLNs and retinas were harvested from the normal diet healthy control (NDC), ketogenic diet heathy control (KDC), normal diet EAU (NDE) and ketogenic diet EAU (KDE) group. Samples were processed via scRNA-seq by using the 10 × Genomics platform. IRBP1-20: Interphotoreceptor retinoid-binding protein 1–20. d Uniform manifold approximation and projection (UMAP) plot: the left showing the cluster of cell types, the right showing sample distribution across two groups. Neu, neutrophils; Macro, macrophages; Mono, monocytes; cDC, classical dendritic cells; pDC, plasmacytoid dendritic cells; NK, natural killer cells. e Pie charts chart showing the percentages of the immune cells derived from scRNA-seq data. f Number of DEGs between KDC and NDC group within each cluster projected onto the UMAP plot. g Volcano plot showing upregulated and downregulated DEGs of total immune cells in the KDC/NDC comparison group. h-i GO terms and KEGG pathways enriched in upregulated (H) and downregulated (I) DEGs of total immune cells in the KDC/NDC comparison group. Significance was calculated based on the accumulative hypergeometric distribution by Metascape webtool
EAU model was induced by the uveitogenic retinal protein, IRBP1–20. On day 14 after immunization, we conducted scRNA-seq on the immune cells from CDLNs of four groups: normal diet healthy control (NDC), ketogenic diet heathy control (KDC), normal diet EAU (NDE) and ketogenic diet EAU (KDE) (Fig. 1c). We firstly explored the impact of KD on healthy mice. Employing classical lineage markers [25,26,27], we identified eight major immune cell types, including T cells, B cells, neutrophils (Neu), monocytes (Mono), macrophages (Macro), classical dendritic cells (cDC), plasmacytoid dendritic cells (pDC) and natural killer cells (NK) (Fig. 1d; Fig. S1a-b). The proportion of T cells showed reduction when transitioning to a KD (Fig. 1e).
To identify the gene signature alterations caused by KD, we performed a DEG analysis. Initially, we quantified the numbers of DEGs, noting their predominance in T and B cells (Fig. 1f). Subsequently, we explored the regulation of specific gene expression. KD was found to downregulate inflammation-related genes (Fos and Junb, both of the AP-1 family) [28], autoimmune disease-related genes (Cebpb and Id2) [29] and lymphocyte activation-related genes (Cxcr4, Il7r, and CD69) [30]. Meanwhile, genes related to Stat1/IFN-γ pathway inhibition (Mbd2) [31], apoptosis (Mcl1) [32] and T cell stemness (Foxo1) [33] were upregulated (Fig. 1g). When compared to the NDC, KDC group exhibited a lower glycolysis score and a higher AMPK signaling pathway score (Fig. S1c-d). In addition, the TGF-β signaling pathway score was increased across most cell subtypes in the KDC (Fig. S1e). Gene function analysis of all immune cells revealed that pathways related to the negative regulation of intracellular signal transduction, histone modification, cellular senescence and FOXO-mediated transcription were upregulated in response to KD (Fig. 1h); and pathways related to cellular responses to stress, TCR signaling and IL-17 signaling were downregulated (Fig. 1i). When attributed to each cell subset, pathways related to mechanisms associated with pluripotency, IL-2 signaling were upregulated (Fig. S1f) across a broad range of cell types; and pathways of neurodegeneration, cellular responses to stress, leukocyte activation, lymphocyte activation were downregulated (Fig. S1g). Overall, these findings suggested that KD may suppress the inflammation- or immune activation-related genes and pathways at baseline, particularly in T cells, highlighting its potential therapeutic benefits.
Ketogenic diet triggers functional changes of T cell and promotes Treg differentiationGiven the aforementioned findings that emphasized the impact of KD on T cells and the well-documented pivotal role of T cells in uveitis, we further re-clustered T cells into nine subsets, including naïve CD4+ T cells (NCD4), regulatory T cells (Treg), T helper 17 cells (Th17), proliferative T cells (ProT), T helper 1 cells (Th1), T follicular helper cells (Tfh), γδT cells (γδT), naïve CD8+ T cells (NCD8) and CD8+ T cells with cytotoxic activity (CTL) (Fig. 2a, Fig. S2a-b). The DEG numbers caused by KD were predominant in Treg, CTL and NCD4 (Fig. 2b), and the upregulated genes was exclusively most in Treg cells (Fig. S2c), including genes that maintain stability (Nrp1) [34], promote cell differentiation (Maf, Orai1) [35, 36] and Treg inhibitory effects (Pvt1) [37] (Fig. 2c). Gene function analysis of Treg cells unveiled several inhibitory pathways were upregulated in KDC group, such as T cell receptor signaling pathway, signaling by TGF−beta receptor complex, FOXO−mediated transcription, and CTLA4 inhibitory signaling (Fig. 2d). Meanwhile, genes specifically downregulated in ProT cells accounted for the largest proportion (Fig. S3a), including cell cycle regulatory genes (Ccnb2, Ccnd2, Cdk4, Cenpa, Mcm7 and Psmd7) (Fid. S3B). The corresponding pathways related to cell cycle were also downregulated (Fig. S3c). These results suggested KD may enhance inhibition function of Treg, while inhibit the proliferation of T cells.
Fig. 2
Ketogenic diet triggered functional changes of T cell and promoted Treg differentiation. a UMAP plot of T cell subsets: the left showing the clusters of all T cell, the right showing sample distribution across KDC and NDC groups. b Wind rose diagram showing the numbers of upregulated and downregulated DEGs in the KDC/NDC comparison group. c Violin plots showing the expression levels of Nrp1, Maf, Pvt1, Orai1 in T cells. d GO terms and KEGG pathways enriched in upregulated DEGs of Treg cells in the KDC/NDC comparison group. Significance was calculated based on the accumulative hypergeometric distribution by Metascape webtool. e Pseudotime trajectory analysis of CD4+ T cells. Cells are colored by pseudotime (left), celltype (middle) and state (right) as indicated. Teff: Th17, Th1, Tfh. f Bar charts showing the percentages of different states between NDC and KDC groups, based on the pseudotime analysis. g-i Proportions of Th17 cells (G), Th1 cells (H) and Treg cells (I) in CDLNs measured by flow cytometry, gated on CD4+T cells. Data expressed as mean ± SD. Significance was determined using unpaired two-tailed student’s t test. ns P > 0.05, ***P < 0.001. j The Treg differentiation rates of NCD4 between NDC and KDC group, measured by flow cytometry, gated on CD4+T cells. Data expressed as mean ± SD. Significance was determined using one-way ANOVA. ****P < 0.0001. NCD4: naïve CD4+ T cells, Treg: regulatory T cells, Th17: T helper 17 cells, ProT: proliferative T cells, Th1: T helper 1 cells, Tfh: T follicular helper cells, γδT: γδT cells, NCD8: naïve CD8+ T cells, CTL: CD8+ T cells with cytotoxic activity
Next, we performed trajectory analysis to clarify the developmental relationships of CD4+ T cells during KD. Ordering of cells in pseudotime arranged most of NCD4, Teff (Th17, Th1 and Tfh) and Treg into an initial trajectory (NCD4-branch) and two bifurcations (Teff-cell fate and Treg-cell fate), respectively. We observed a pseudotemporal path started from NCD4 to Teff and Treg (Fig. 2e). Compared to the NDC group, the KDC group showed an increased ratio of Treg-cell fate, with a merely unchanged ratio of Teff-cell fate (Fig. 2f). By flow cytometry, KD didn’t influence the proportions of Th1 and Th17 cells, whereas significantly increase the Treg cells (Fig. 2g–i), indicating that KD may promote Treg differentiation. To validate this hypothesis, NCD4 cells were isolated from both NDC and KDC mice, and then cultured under Treg-polarizing conditions for 72 h. Notably, the KDC group displayed a higher rate of Treg differentiation (Fig. 2j). Collectively, these findings suggest that KD treatment can promote Treg differentiation.
Ketogenic diet induces changes in B cell transcriptional profile and immune cell–cell communicationsFor a comprehensive illustration of KD’s impact, we further re-clustered B cells into three subsets, including naïve B cells (NBC), germinal B cells (GBC), plasma B cells (PBC) (Fig. 3a, Fig. S4a-b). By SCENIC (Single Cell Regulatory Network Inference and Clustering), the transcription factor (TF) regulatory networks were predicted to determine how KD regulates B cell transcriptional profiles. Among the top 10 TFs of the two group: autoimmune related regulator, Junb and Cebpb, showed decreased transcriptional activity in KDC mice, while Bcl11a and Rfx7 ( DNA repair and anti-apoptosis regulatory molecule [38, 39]) displayed elevation (Fig. 3b). In addition, DEG analysis of NBCs revealed that genes associated with inflammation (Cd24a, Cxcr4, Id2, Cd69 and Cebpb) [40], MAPK signaling pathway (Junb, Fos, Dusp1), glycolysis regulatory (Myc) [41] and immunoglobulin (Igkc, Ighm) were downregulated in the KDC group (Fig. 3c). Pathways related to T or B cell activation, differentiation, and signaling were downregulated (Fig. 3d).
Fig. 3
Ketogenic diet induced changes in B cell transcriptional profile and immune cell–cell communications. a UMAP plot of B cell subsets: germinal center B cells (GBC), naïve B cells (NBC), and plasma cells (PC). b Heatmap exhibiting the average regulon activities of top 10 transcription factors in KDC and NDC groups, color key from white to red indicates relative expression levels from low to high. c Volcano plot showing upregulated and downregulated DEGs of NBC in the KDC/NDC comparison groups. Significance was determined using “FindMarkers” functions of Seurat package with Wilcoxon Rank Sum test and adjusted by Bonferroni correction. d Downregulated GO terms and KEGG pathways enriched in NBC cells in the KDC/NDC comparison group. Significance was calculated based on the accumulative hypergeometric distribution by Metascape webtool. e–g Circle plot showing the TGF-β signaling (e), CXCL signaling (f), ICAM signaling (g) of immune cell subsets in KDC and NDC groups, calculated by CellChat. The thickness of the lines represents the numbers of interaction. h Dotplot showing the relative expression of interaction pairs from cDC to γδT cells (left) and Th17 cells (right), respectively
Secondly, we compared the relative regulon activity of myeloid cells: the regulators related to autoimmune disease (Irf5), oxidative stress (Klf9) and macrophage activity (Bhlhe40) were decreased in KDC group (Fig. S4c). DEG numbers were exclusively most in cDC cells, whatever upregulated or downregulated in KDC group (Fig. S4d). Therefore, we performed gene function analysis of cDC cells. The downregulated DEGs in KDC were enriched in pathways related to neutrophil degranulation, antigen processing and presentation and mononuclear cell migration, while the upregulated are related to negative regulation of immune system process and leukocyte cell−cell adhesion (Fig. S4e, S5a). Together, these results suggested that KD may inhibit inflammatory activation of B cells and the immune response of cDC cells.
Lastly, using the CellChat, we compared signaling pathway networks among all immune cell subtypes between the KDC and NDC groups. The circle plots showed that the TGF-β signaling pathways were enhanced by KD, and the branches of endpoint to Treg cells were enriched but absent in the NDC group (Fig. 3e). The CXCL signaling pathways (from cDC cells to Th17 and γδT cells) and ICAM signaling pathways (from cDC cells to effector CD4+ T and ProT cells) were enriched in NDC group but decreased in KDC group (Fig. 3f–g). Furthermore, SCENIC analysis validated that the chemotaxis signaling pathways between cDC and Th17, γδT cells were indeed weakened (Fig. 3h; Fig. S5b). As dendritic cells are crucial antigen presented cells, the chemotaxis signaling pathways associated with inflammation were also decreased between pDC and all T cell subtypes (Fig. S5c–e). These results indicated that KD could weaken the inflammatory cell–cell communications.
Ketogenic diet alleviates EAU inflammation and restores the Th17/Treg balanceTo further explore the effect of KD on autoimmune diseases, we next conducted experiments on EAU, a classical animal model of uveitis. The KDE group displayed mild inflammatory infiltrate with markedly reduced clinical scores (Fig. 4a). Histopathological manifestations aligned with clinical observations (Fig. 4b), in that the KDE group showed only slight inflammatory cell infiltrations. The scRNA-seq analysis revealed a reduction of T cell ratio in the KDE (Fig. 4c).
Fig. 4
Ketogenic diet alleviated EAU inflammation and restored the Th17/Treg balance. a Representative fundus images and clinical scores of eyes from the NDC, NDE and KDE group, captured on day 14 post-immunization through fundus photography. Red arrows indicate inflammatory exudation and vascular deformation. Data represented as mean ± SD. Significance was determined using one-way ANOVA. ***P < 0.001, ****P < 0.0001. b Representative histopathological images (hematoxylin and eosin staining) and pathological scores of retinas from the NDC, NDE and KDE group, captured on day 14 post-immunization. Red arrows indicate infiltration of inflammatory cells and retinal folding. Data represented as mean ± SD. Significance was determined using one-way ANOVA. ****P < 0.0001. c Pie charts showing the percentages of the immune cell subsets in CDLNs from NDE and KDE groups. d Wind rose diagrams showing the numbers of upregulated and downregulated DEGs in the KDC/NDC and KDE/NDE comparison groups. e Wind rose diagrams showing the numbers of upregulated and downregulated DEGs in the NDE/NDC and KDE/KDC comparison groups. f Pie charts showing the percentages of the T cell subsets in CDLNs from NDE and KDE groups. G Violin plots showing expression of Il17a and Foxp3 in CD4+T cells in NDE and KDE groups. h-i Proportions of Th17 cells (h), Th1 cells (i) and Treg cells (j) in CDLN CD4+T cells measured by flow cytometry on day 14 post-immunization from the NDC, NDE and KDE group. Data expressed as mean ± SD. Significance was determined using one-way ANOVA. ns P > 0.05, *P < 0.05, **P < 0.01, ***P < 0.001
Subsequently, we explored its regulation on specific gene expression. Compared to the ND group, the DEG numbers of KDC and KDE group were consistently concentrated in T cells and B cells (Fig. 4d). When compared to healthy control group, the NDE exhibited a high number of DEGs in T and B cells, whereas the KDE showed a low number (Fig. 4e). These downregulated genes in the KDE group were related to inflammation (Cxcr4, Atf4 and Cebpb), MAPK signaling pathway (Junb, Fos, Fosb, Jund) and Th17 cell activation (Pim1) (Fig. S6a). Gene function analysis of all immune cells revealed that pathways associated with cellular responses to stress, T cell activation and IL-17 signaling were downregulated in KDE group, while negative regulation of immune system process and PD−1 checkpoint pathways were upregulated (Fig. S6b-c). We also found that the IL-17 pathway was downregulated in almost all immune cell subtypes, while the AMPK pathway was upregulated (Fig. S6d). These results indicated that KD rescued the changes in gene expression caused by autoimmune responses.
Since autoimmune diseases, including uveitis, are often characterized by the aberrant differentiation of CD4+ T cells [3, 42,43,44], we subsequently focused on these cell subsets. Based on scRNA-seq, the proportion of Th17 and Th1 cells was declined and the Treg was increased in the KDE group (Fig. 4f), consistent with the average expression levels of Il17a and Foxp3 in CD4+ T cells (Fig. 4g). Flow cytometry results further confirmed these findings, and the Th1 showed a declined tendency (Fig. 4h-j). Changes in splenic CD4+T cell subtypes showed a consistent trend (Fig. S6e-g). Further studies found that the expression levels of T cell activation and inflammation related genes were decreased in almost all T cell subsets, especially in effector CD4+ T cells (Fig. S7a). Pathways related to IL−17 signaling pathway and cell cycle were downregulated in Th17 cells, while T cell activation and proliferation were upregulated in Treg cells (Fig. S7b-c). Importantly, when CD4+ T cells from the NDE and KDE mice were transferred into blank mice, respectively, the KDE group exhibited attenuated pathogenicity (Fig. S7d-e). These results demonstrated that KD effectively alleviated EAU progression and restored Th17/Treg balance.
Ketogenic diet inhibites immune cell infiltration and inflammatory response in retinaRetinal damage is the main clinical manifestation in uveitis. Therefore, we sorted the retinal immune cells for scRNA-seq to elucidate the effect of KD on local immune microenvironment. We clustered immune cells infiltrating in the retina into eight subsets, including T cells, B cells, neutrophils (Neu), monocytes and macrophages (MM), classical dendritic cells (cDC), plasmacytoid dendritic cells (pDC), microglias (MiG) and natural killer cells (NK) (Fig. 5a; Fig. S8a). The proportion of retina-infiltrating T cells also decreased after the KD (Fig. 5b). The expression of IL17 signaling-related genes (Id2, Cd69 and Pim1) [40] and glycolysis regulatory gene (Hif1α) [41] were downregulated in response to KD, while genes involved in retinal development were upregulated (Gnb1, Prph2, Pde6b, Prkn) (Fig. 5c). Gene function analysis of all immune cells unveiled pathways related to regulation of cell activation, TCR signaling were downregulated; while pathways associated with negative regulation of intracellular signal transduction, myeloid cell and neutrophil apoptotic process were upregulated (Fig. S8b-c). Further subtype analysis demonstrated a significantly lower Th17 cell differentiation score in T cell and Neu subsets of the KDE group (Fig. 5d), accompanied by increased AMPK signaling pathway scores in most subtypes (Fig. S8d).
Fig. 5
Ketogenic diet inhibited immune cell infiltration and inflammatory response in retina. a UMAP plot clustering of retinal immune cells from NDE and KDE groups. b Bar chart showing the percentages of immune cell subsets in the NDE and KDE groups derived from scRNA-seq data. c Volcano plot showing upregulated and downregulated DEGs of all retinal infiltrating immune cells in the KDE/NDE comparison groups. Significance was determined using “FindMarkers” functions of Seurat package with Wilcoxon Rank Sum test and adjusted by Bonferroni correction. d Box plot showing Th17 cell differentiation scores across retinal immune subsets from NDE and KDE groups. e UMAP plot clustering of T cells from NDE and KDE groups. f Bar chart showing the percentages of T cell subsets in the NDE and KDE groups derived from scRNA-seq data. g-j Proportions of Th17 cells (g), Th1 cells (h), Treg cells (i) in retinal CD4+T cells, and CD4+T cells (j) measured by flow cytometry on day 14 post-immunization from the NDE and KDE group. Data expressed as mean ± SD. Significance was determined using one-way ANOVA. ns P > 0.05, **P < 0.01, ****P < 0.0001. k–l Circle plot showing MHC-II signaling (k), TGF-β signaling (l) of immune cell subsets in NDE and KDE groups, calculated by CellChat. The thickness of the lines connecting cells indicates the numbers of interaction. MM: monocytes & macrophages, MiG: microglias
Based on scRNA-seq, the proportion of CD4+T, Th17 and Th1 cells exhibited a decreased trend in the KDE group (Fig. S8f, Fig. 5f). The inflammation-related genes (B2m and Pim1) [40] and glycolysis regulatory gene (Hif1α) [41] were downregulated in KDE group (Fig. S8g). The pathways related to necroptotic cell death, retinal cell apoptotic process, regulation of protein catabolic process were upregulated in KDE group; while regulation of T cell activation, Th17 cell differentiation, PIP3 activates AKT signaling and TCR signaling were downregulated (Fig. S8h-i). Flow cytometry results showed that in the KDE group, the proportions of CD4+ T cells and Th17 cells infiltrating in the retina were significantly reduced, the Treg cells was significantly increased, and the Th1 cells showed the declined trend (Fig. 5g–j). Additionally, using the CellChat, we compared the cell–cell communication between the KDE and NDE groups. The circle plots showed that the MHC-II signaling pathways were weakened in the KDE group, while the TGF-β signaling pathways were enhanced (Fig. 5k–l). Collectively, these results suggested that KD could prevent the immune cell infiltration into the retina and mitigate the inflammatory response therein.
Integrated analysis of immune cell subsets from retina and CDLNs in KD miceIn uveitis, pathogenic immune cells in LNs migrate to the retina, altering retinal immune microenvironment [27] and causing damage. Therefore, we integrated the scRNA-seq data from retina and CDLNs (Fig. S9a), revealing significant heterogeneity (Fig. 6a). In terms of proportions, B and T cells dominated in CDLNs; while cDC, Microglia, MM, Neu and NK were more abundant in retina (Fig. 6b; Fig. S9b-c). In addition, KD simultaneously decreased the ratio of NK and T cells in retina and CDLNs. Whether in CDLNs or retina, genes related to T cell activation (Cd69, Rac1), IL17 pathway (Il1b, Cebpb), and autoimmune response (Bhlhe40, Fos, S100a10, S100a13, S100a4, S100a6) were downregulated in the KDE group, and most of them express higher levels in the retina (Fig. 6c; Fig. S9d). Multiple immune-inflammatory pathways were upregulated from LN to retina, but exhibited a mitigated tendency in the KDE group (Fig. 6d), which was same goes for immune cell subtype (Fig. S10a).
Fig. 6
Merged analysis of immune cell subsets from retina and CDLNs in KD mice. a UMAP plot clustering of retina and CDLN immune cells from NDE and KDE groups. b Bar chart showing the percentages of immune cell subsets form retina and CDLN of NDE and KDE groups derived from scRNA-seq data. c Line charts showing the mean expression of Pim1, Cxcr4, Hif1a, Id2 in retina and CDLN immune cells from NDE and KDE groups. d Representative GO terms and pathways enriched in upregulated DEGs in retina/LN comparison group between NDE and KDE groups. e UMAP plot clustering of retina and CDLN T cells from NDE and KDE groups. f-g Pseudotime trajectory analysis of CD4+ T cells from retina and CDLN of NDE and KDE groups. Cells are colored by pseudotime (right), celltype (left) as indicated (f). Cells are colored by celltype, splited by tissues and group as indicated (g). h–k The scatter distribution plot showed the expression alteration of Pim1, Cxcr4, Ilb, Id2 in each group during the pseudotime. The color coded for different tissues
To delineate the KD’s impact on evolution tendency of CD4+ T cell from CDLNs to retina, we further re-clustered the merged T cell subsets (Fig. 6e) and conducted a trajectory analysis using monocle 2. The pseudotime ordering of cells were started from NCD4 to Th1_17_fh (Th17, Th1 and Tfh) and Treg cells (Fig. 6f). The CDLN CD4+ T cells were mainly located at the starting pseudotime, while the retinal predominantly accumulated at the ending pseudotime, and almost no NCD4 in the retina (Fig. 6g). In addition, the retinal CD4+ T cells are at a higher degree of developmental status, and KD restrain the development of Th1_17_fh (Fig. 6g). Analysis of cell ratios revealed that state 1 was dominated by NCD4 cells, both state 2 and state 3 were dominated by Th1_17_fh fate cells, with state 3 containing more Treg cells (Fig. S10b). In the KDE group, the proportion of NCD4 cells in state 1 and the Treg cells in state 3 were both upregulated, while the Th1_17_fh in state 3 was declined (Fig. S10b). Moreover, KD suppressed the expression of genes related to immunological pathways (Cxcr4, Ilb2) and IL-17 signaling (Pim1, Id2) with pseudotime (Fig. 6h–k). Together, these results suggested that retinal immune cells exhibited greater inflammatory responses than CDLN cells in uveitis, which KD could partially counteract.
留言 (0)