
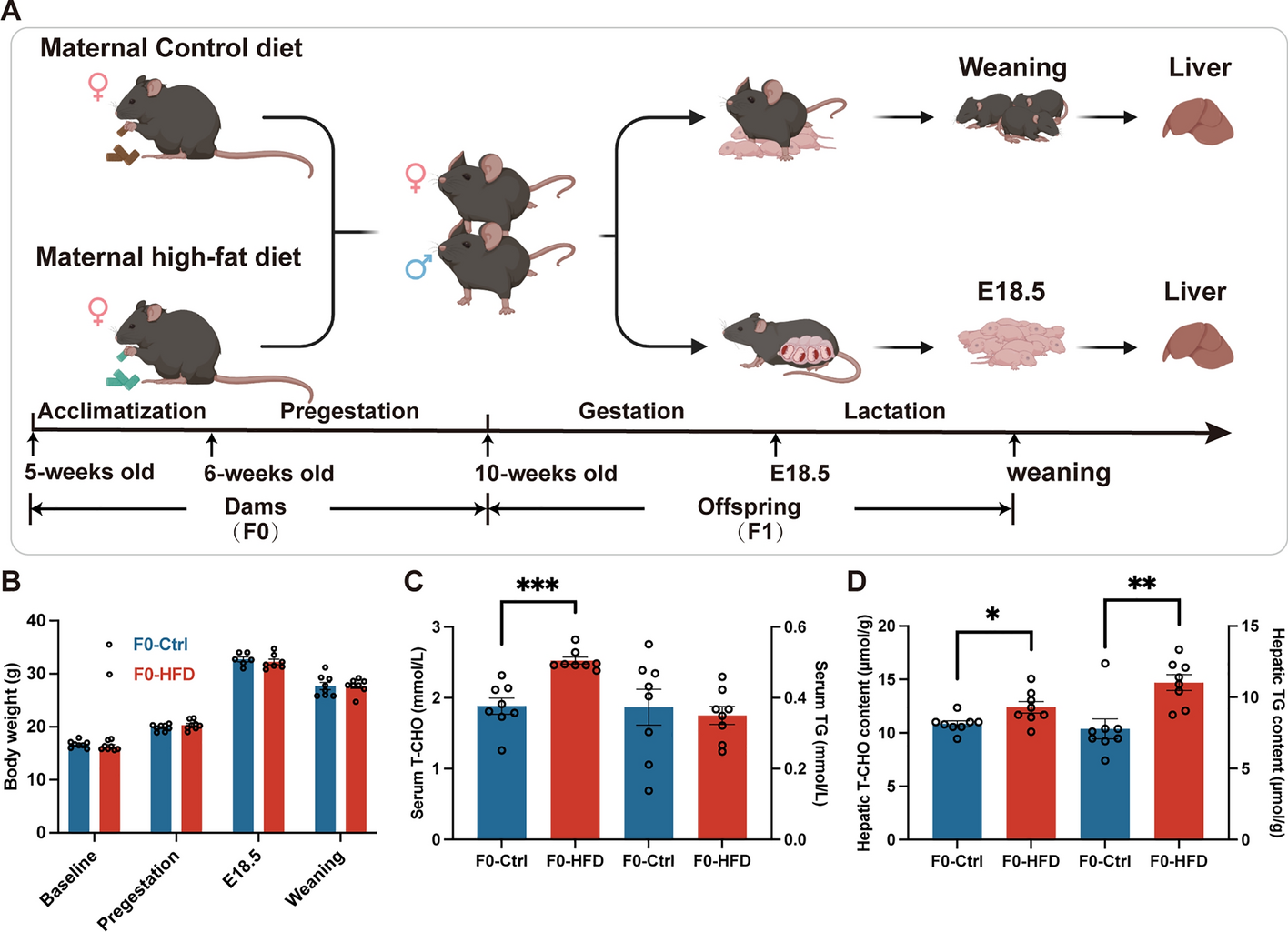
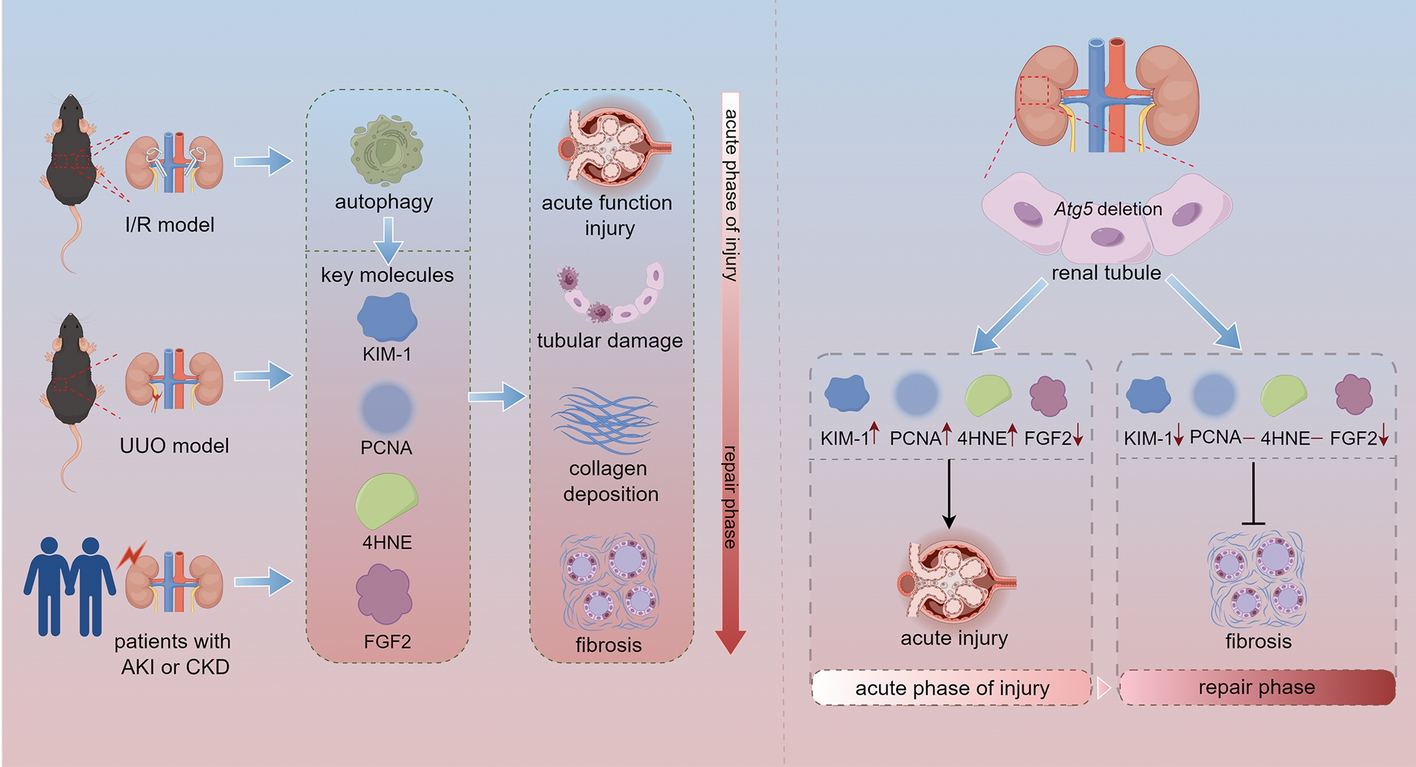
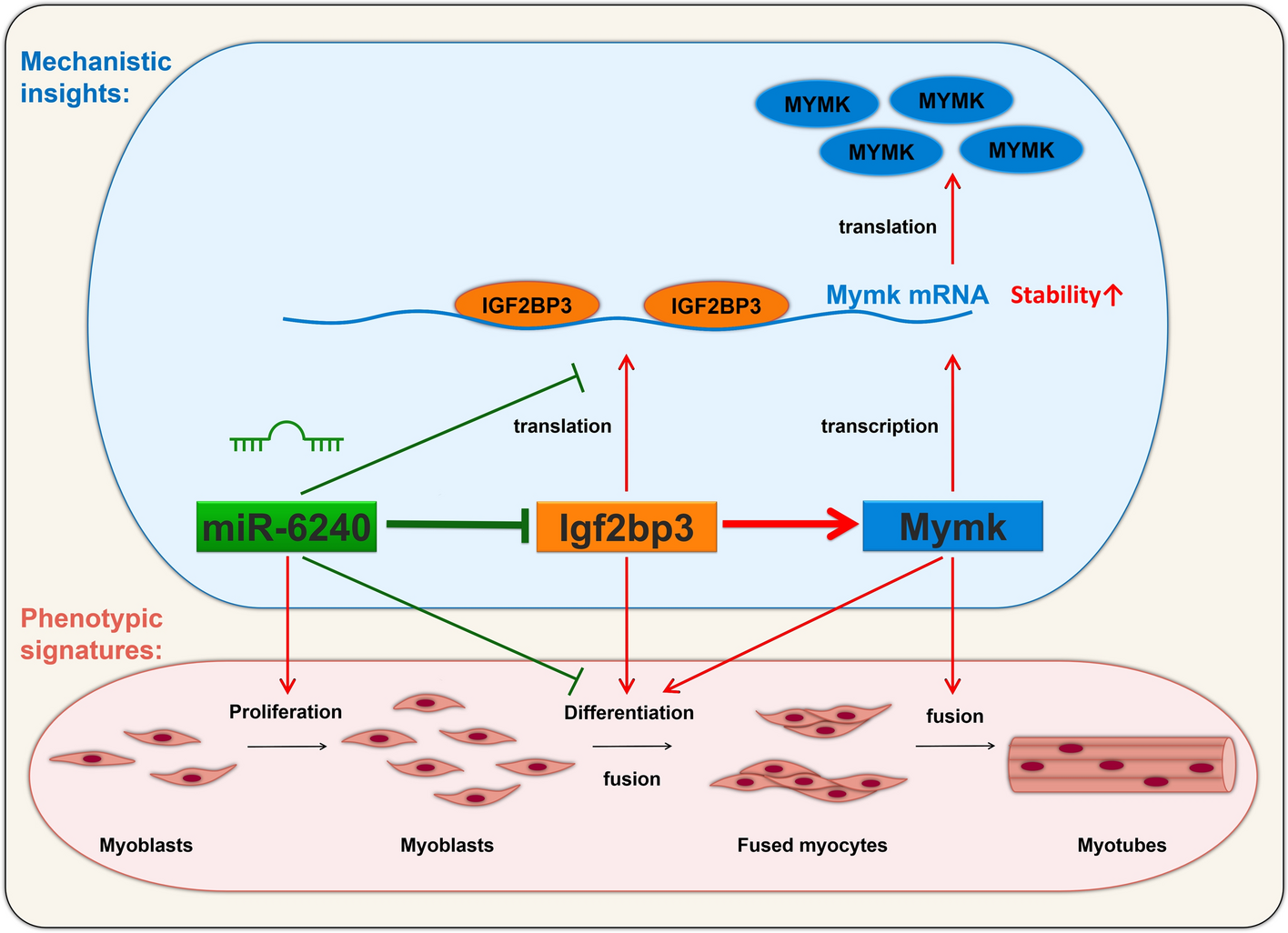
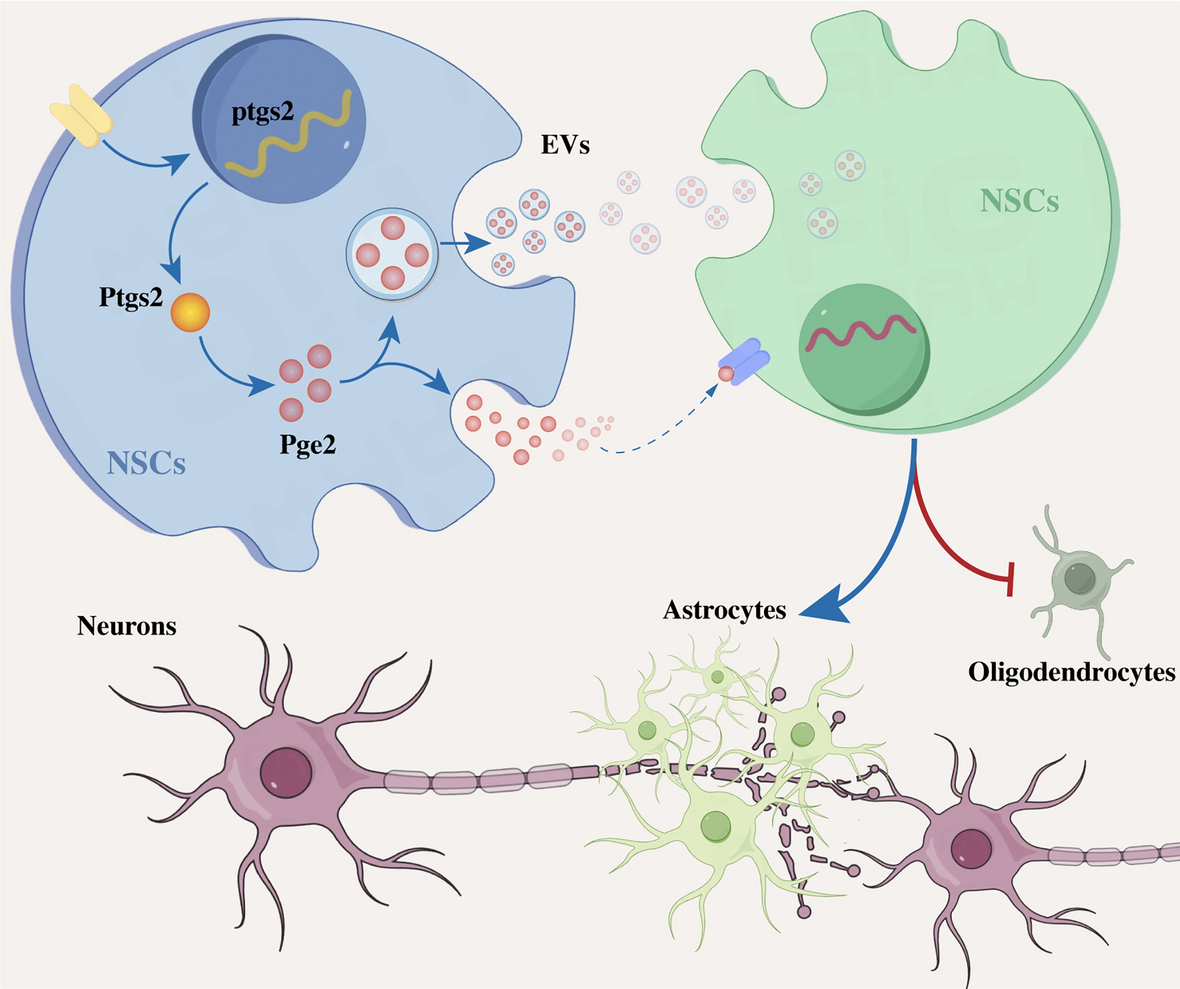
記住我



To assess the expression of PCDHGA9 in colorectal cancer tissues, we selected 63 paired tissue specimens from colorectal cancer patients who had undergone radical surgery at Shanghai General Hospital between 2013 and 2014, which were then subjected to RNA extraction and subsequent qRT-PCR analysis. The qRT-PCR results indicated that, among the 63 colorectal cancer patient tissues, 42 patients exhibited lower mRNA expression of PCDHGA9 in CRC tissue compared with paracancerous tissue (Fig. 1a), and overall survival and disease-free survival analysis demonstrated that colorectal cancer patients with reduced PCDHGA9 expression had poorer survival rates (Fig. 1b, c). Furthermore, poorly differentiated CRC tissues exhibited lower expression of PCDHGA9 (Fig. 1e).
Fig. 1
Comprehensive investigation of PCDHGA9 in CRC: expression in tissue, survival analysis, cell line manipulation, and high-throughput sequencing results. a Real-time quantitative PCR (qPCR) revealed decreased PCDHGA9 expression in 42 CRC tissues among 63 paired samples, compared with adjacent normal tissues. b, c Patients with low PCDHGA9 expression exhibited shorter overall survival and disease-free survival rates. d, f Western blot analysis assessed PCDHGA9 expression in HCT116, RKO, HCT8, and SW620 cells, as well as the impact of overexpressing or knocking down PCDHGA9 on PCDHGA9 and β-catenin expression. e Immunohistochemistry exhibits the expression of PCDHGA9 in CRC tissues with low, moderate, and high differentiation. g High-throughput sequencing identified differential gene expression between vector and PCDHGA9-overexpressing cells, with downregulated genes listed on the right
To further investigate the significance of PCDHGA9 in colorectal cancer, we collected clinicopathological information of CRC patients, which was then assessed in combination with PCDHGA9 expression, particularly in regards to patient age, gender, tumor location, tumor size, tumor stage, tumor differentiation, tumor invasion degree, and lymphatic vascular metastasis. The chi-squared test showed that patient age, gender, tumor location, and tumor size were not related to PCDHG9 expression, while significant association was observed with patient tumor stage, tumor differentiation, tumor invasion degree, and lymphatic vascular metastasis (Table 4). Taken together, on the basis that these tumor characteristics, namely tumor stage, tumor differentiation, extent of tumor invasion, and lymphatic vascular metastasis, closely correspond to the inherent invasive and metastatic attributes of tumor cells, we speculated a correlation between PCDHGA9 and tumor invasion and metastasis.
Table 4 Correlation between PCDHGA9 expression and clinical information of patients (n = 63)Downregulated PCDHGA9 promotes invasion, metastasis, and EMT of CRC cellsTo verify this speculation, we detected the mRNA and protein expression of PCDHGA9 in four cell lines: HCT8, HCT116, RKO, and SW620. The results showed a relatively lower PCDHGA9 expression in HCT116 and RKO cells and higher expression in HCT8 and SW620 cells (Fig. 1d, Supplementary Fig. S1a). Subsequently, we employed lentiviral technology to construct an overexpressing PCDHGA9 cell line in HCT116 and RKO and a lentiviral vector containing short hairpin RNA targeting PCDHGA9 cell line in HCT8 and SW620 (Fig. 1f, Supplementary Fig. S1bc). Differential gene analysis was conducted between vector of PCDHGA9 and overexpression of PCDHGA9 in HCT116 using high-throughput sequencing (Fig. 1g). Clustering analysis of the differential genes using ingenuity pathway analysis (IPA) unveiled an association between PCDHGA9 and cell adhesion function (Supplementary Fig. S1e).
Both the analysis of pathological data and sequencing results have confirmed a correlation between PCDHGA9 expression and the invasive and metastatic functions of CRC. Subsequently, we investigated the impact of PCDHGA9 on the invasion and metastasis abilities, along with the epithelial–mesenchymal transition (EMT) of CRC cells, through in vitro cellular assays. Wound healing and Transwell assays were performed to examine the influence on CRC cells’ migration and invasion abilities, and the results showed that knockdown of PCDHGA9 led to a faster scratch healing rate among CRC cells and an increased number of cells invading through the Transwell membrane compared with the negative control (NC) groups, while opposite results were observed in the overexpression of PCDHGA9 groups (Fig. 2a, b). Experiments involving assessment of the expression of EMT-related proteins showed that the knockdown of PCDHGA9 resulted in a significant decrease in E-cadherin levels and a significant increase in N-cadherin, vimentin, and Snail expression, while the overexpression of PCDHGA9 resulted in a significant increase in E-cadherin expression and significant reductions in N-cadherin, vimentin, and Snail levels (Fig. 2c).
Fig. 2
PCDHGA9 suppresses invasion, metastasis, and EMT of CRC cells. a Wound healing assays were conducted on CRC cells with overexpressed or downregulated PCDHGA9 and their respective controls. The width of wounds was measured at 0 and 48 h, and the percentage of wound closure was calculated and analyzed using the Student’s t-test. b Transwell assays were performed on CRC cells with manipulated PCDHGA9 expression and their controls, and the number of cells that had migrated through the chamber was observed and quantified. c Western blot analysis showing changes in E-cadherin, N-cadherin, vimentin, and Snail expression following the overexpression or downregulation of PCDHGA9. Notably, E-cadherin was not detected in RKO cells from our laboratory, resulting in the absence of an E-cadherin blot
Our previous investigation in gastric cancer revealed the inhibitory role of PCDHGA9 in the Wnt/β-catenin pathway, and here we extended similar research in colorectal cancer. We performed western blot (WB) to assess β-catenin protein levels and found a significant reduction in the intracellular β-catenin protein content in HCT116 and RKO cells following PCDHGA9 overexpression compared with the vector, and this pattern was reversed when PCDHGA9 was knocked down in HCT8 and SW620 cells (Fig. 1g). On the basis of these outcomes, we speculated that PCDHGA9 potentially reduces the invasion and metastasis of CRC by facilitating the degradation of β-catenin, thereby leading to a reduction in β-catenin protein present in CRC cells.
miR-1269a downregulates PCDHGA9 and promotes invasion, metastasis, and EMT of CRC cellsMicroRNAs have attracted substantial attention in recent tumor research owing to their pivotal roles, with numerous studies showing the association between dysregulation of oncogenes and microRNAs [23,24,25]. To further investigate the underlying cause for the reduced PCDHGA9 expression in colorectal tumors, we conducted a comprehensive analysis to predict the upstream regulatory microRNAs of PCDHGA9 using the StarBase, TargetScan, and miRBD bioinformatics tools, which identified hsa-miR-1269a and hsa-miR-3179 as common target microRNAs predicted by all three tools (Fig. 3a). However, our literature search showed that hsa-miR-5586-5p, hsa-miR-580-3p, hsa-miR-760, and hsa-miR-625-5p, which were identified as target microRNAs, demonstrated low expression in tumors or acted as inhibitors of cancer progression in previous studies [26,27,28,29]. This inconsistency with the anticipated goal of identifying highly expressed microRNAs that contribute to reduced PCDHGA9 expression and promote CRC progression prompted us to focus on miR-1269a and miR-3179. Then, we transfected miR-1269a and miR-3179 mimics into 293T cells, which revealed a robust downregulatory effect of miR-1269a on PCDHGA9, while miR-3179 exhibited a comparatively weaker effect on the downregulation of PCDHGA9 (Fig. 3b). Consequently, we selected miR-1269a as a potential upstream regulator of PCDHGA9 in the subsequent experiments.
Fig. 3
miR-1269a targets PCDHGA9 and downregulates PCDHGA9. a Venn diagram illustrating the microRNAs predicted to interact with PCDHGA9 across three databases (starBase, Targetscan, and miRBD). Among these, hsa-miR-1269a, hsa-miR-5586-5p, hsa-miR-580-3p, hsa-miR-760, hsa-miR-625-5p, and hsa-miR-3179 were identified as commonly predicted microRNAs. b PCDHGA9 expression was assessed via qPCR after transfection of miR-1269a and miR-3179 mimics into 293T cells. c Four groups of 293T cells were transfected with different combinations of vectors (NC or miR-1269a mimic) and luciferase reporter plasmids (wild type or mutant type PCDHGA9). The luciferase activity was measured using the dual luciferase assay system. d The relative expression of PCDHGA9 was examined by qPCR following the transfection of miR-1269a inhibitor or mimic into CRC cells. e The protein levels of PCDHGA9 and β-catenin were analyzed by western blotting in CRC cells transfected with miR-1269a inhibitor or mimic. f The interacting region between PCDHGA9 and miR-1269a is shown
HCT8 and SW620 cells were transfected with the miR-1269a mimic, whereas HCT116 and RKO cells were transfected with the miR-1269a inhibitor (Supplementary Fig. S2ab). Subsequently, we assessed the RNA and protein expression of PCDHGA9, revealing that miR-1269a exhibited a downregulatory effect on PCDHGA9 (Fig. 3d, e). To further validate the regulatory relationship between miR-1269a and PCDHGA9, luciferase assays were conducted in 293T cells. The results demonstrated that the miR-1269a mimic could effectively downregulate the luciferase expression of wild-type PCDHGA9 transcripts, which was nullified upon mutation of the predicted binding site, emphasizing the specificity of the interaction (Fig. 3c, f). These findings collectively confirm that miR-1269a exerts its influence by downregulating PCDHGA9 through the predicted binding site.
Next, wound healing and Transwell assays were performed to detect the effects of miR-1269a on CRC cells’ invasion and metastasis ability, and the results showed that miR-1269a promoted the invasion and metastasis of CRC cells (Fig. 4a, b). Western blot analysis was conducted to evaluate EMT-related proteins and β-catenin expression, which showed that miR-1269a not only promoted the process of EMT in CRC cells but also led to an increase in β-catenin levels (Figs. 3e, 4c).
Fig. 4
MiR-1269a promotes invasion, metastasis, and EMT of CRC cells. a Wound healing assays were conducted in CRC cells transfected with either miR-1269a inhibitor or mimic, along with their respective controls. The widths of wounds were measured at 0 h and 48 h, and the percentages of wound closure were calculated and analyzed. b Transwell assays were performed in CRC cells transfected with either miR-1269a inhibitor or mimic, along with their respective controls. The cells that had migrated out of the chamber were observed and counted. c Western blotting results demonstrated changes in the protein levels of E-cadherin, N-cadherin, vimentin, and Snail after transfection with either miR-1269a inhibitor or mimics into cells
CXCR4 is downregulated by PCDHGA9 interacting with HOXA1 and affects invasion, metastasis, and EMT of CRC cellsTo further investigate the mechanism of β-catenin regulation by PCDHGA9, high-throughput sequencing results revealed a significant reduction in CXCR4 expression in cells overexpressing PCDHGA9. qRT-PCR analysis corroborated that both PCDHGA9 overexpression and knockdown led to corresponding changes in CXCR4 expression (Supplementary Fig. S2c, d). Considering that numerous studies have highlighted CXCR4’s role in inhibiting β-catenin degradation through the AKT/GSK-3β pathway, triggering intracellular β-catenin accumulation, Wnt pathway activation, and fostering cancer invasion and metastasis [30,31,32], we examined CXCR4 as a potential downstream target of PCDHGA9.
Recognizing that PCDHGA9 functions as a transmembrane protein rather than a direct transcription factor, its capacity for gene transcription is limited. On the basis of the interaction potential of the calmodulin protein family, we explored their influence on intracellular distribution and functionality. Through coimmunoprecipitation assay, we identified coprecipitated proteins in overexpressing PCDHGA9 cells, including transcription factors HOXA1, HOXB3, and HOXB13 (Supplementary Fig. S2e). According to JASPAR prediction, HOXA1 exhibited multiple high-affinity binding sites within the CXCR4 promoter region (Supplementary Fig. S2f), which reveals HOXA1 as an intermediary in the PCDHGA9–CXCR4 nexus. Western blot analysis confirmed PCDHGA9’s interactions with HOXA1, revealing its role in modulating HOXA1 distribution, particularly increasing cytoplasmic levels while reducing nuclear presence (Figs. 5a, 7c, d). The overexpression and knockdown of PCDHGA9 decreased and increased the activity of a luciferase plasmid driven by the CXCR4 promoter sequence, respectively (Fig. 5b). These results show the potential of PCDHGA9 in modulating the distribution of HOXA1, thereby influencing CXCR4 expression through the regulation of CXCR4 transcription.
Fig. 5
CXCR4 is downregulated by PCDHGA9 interacting with HOXA1. a Co-IP assays revealed the presence of the FLAG and HOXA1 proteins in both the input and immunoprecipitated samples. b Dual luciferase assays showing the relative activity of the wild-type (WT) CXCR4 luciferase reporter plasmid after either overexpressing or downregulating PCDHGA9. c Predicted binding sites of HOXA1 within the CXCR4 promoter region at positions 1437, 400, and 77. Two mutant constructs, MUT1 and MUT2, were generated for the three predicted binding sites. Dual luciferase assays were conducted to assess the relative activity of WT, MUT1, and MUT2 CXCR4 luciferase reporter plasmids in the presence or absence of HOXA1. d Western blot analysis depicting the protein expression of CXCR4, PCDHGA9, and β-catenin in CRC cells with altered CXCR4 expression
To confirm HOXA1’s role as a CXCR4 transcription factor, we introduced a plasmid containing the HOXA1 transcript into HCT116 cells, which resulted in increased CXCR4 expression (Fig. 7b). Further, we created two mutations (MUT1 and MUT2) within the three predicted binding sites (Fig. 5c) and introduced them along with vector and HOXA1 overexpression. The results showed that HOXA1 overexpression enhanced CXCR4 expression across the three groups. However, when the CXCR4 promoter was mutated, the increase caused by HOXA1 was reduced (Fig. 5c), demonstrating that HOXA1 acts as a transcription factor for CXCR4, promoting its expression.
Combined with the above experiments, our analysis reveals that PCDHGA9 influences CXCR4 expression by interacting with HOXA1, a transcription factor for CXCR4, and influencing the intracellular distribution of HOXA1.
Next, we examined the impact of CXCR4 on the invasive and metastatic abilities of CRC cells. The results showed that CXCR4 promotes invasion and metastasis (Fig. 6a, b), stimulates EMT, and increases β-catenin levels in CRC cells (Figs. 5d, 6c). Importantly, it was observed that CXCR4 did not influence PCDHGA9 (Fig. 5d).
Fig. 6
CXCR4 promotes invasion, metastasis, and EMT of CRC cells. Wound healing (a), Transwell assays (b), and WB (c) showing the effects of CXCR4 on promoting the invasion, metastasis, and EMT of CRC cells
The regulatory relationship among PCDHGA9, miR-1269a, CXCR4, and β-cateninWe conducted retrospective experiments to confirm the interplay among PCDHGA9, miR-1269a, CXCR4, and β-catenin. Previous assays established that miR-1269a downregulates PCDHGA9, which then downregulates CXCR4 to affect β-catenin levels in CRC cells. However, altering PCDHGA9 expression did not affect miR-1269a levels when overexpressed or knocked down (Supplementary Fig. S1d). Similarly, CXCR4 had no impact on PCDHGA9 or miR-1269a (Figs. 5d, 7a). Then, we assessed their combined impact on downstream factors and found that miR-1269a downregulated both CXCR4 and β-catenin, whereas PCDHGA9 counteracted this downregulation (Fig. 7c). Moreover, miR-1269a counteracted the PCDHGA9-mediated transfer of HOXA1 from the nucleus to the cytoplasm (Fig. 7d). Next, we introduced (E)-ferulic acid, a β-catenin inhibitor, to vector and overexpressing CXCR4 cells and observed that (E)-ferulic acid impeded Wnt/β-catenin pathway activation driven by CXCR4 overexpression without significantly affecting miR-1269a, PCDHGA9, or CXCR4 expression (Fig. 7e, Supplementary Fig. S3a). Similarly, CHIR-99021, a β-catenin activator, was introduced to vector and overexpressing PCDHGA9 cells, which then counteracted the β-catenin reduction induced by PCDHGA9 while showing no substantial effect on miR-1269a, PCDHGA9, or CXCR4 (Fig. 7e, Supplementary Fig. S3b). Collectively, these findings suggest that β-catenin functions as a downstream regulatory target of miR-1269a, PCDHGA9, and CXCR4, which collectively constitute the miR-1269a/PCDHGA9/CXCR4/β-catenin pathway, which can be targeted to prevent CRC invasion and metastasis (Fig. 7f).
Fig. 7
The regulatory relationship among PCDHGA9, miR-1269a, CXCR4, and β-catenin. a Quantitative PCR (qPCR) analysis demonstrated the expression levels of miR-1269a following the downregulation or overexpression of CXCR4. b Western blotting (WB) revealed the protein expression of CXCR4 and β-catenin in the presence or absence of the HOXA1 overexpressing plasmid. c WB showed alterations in the protein levels of HOXA1, CXCR4, and β-catenin upon PCDHGA9 overexpression and the introduction of the miR-1269a mimic. d WB depicted changes in the HOXA1 distribution between the cytoplasm and nucleus in the presence or absence of PCDHGA9 overexpression and the miR-1269a mimic. e WB analysis demonstrated that (E)-ferulic acid, a β-catenin inhibitor, affected β-catenin expression without impacting PCDHGA9 and CXCR4. Similarly, CHIR-99021, a β-catenin activator, influenced β-catenin expression while leaving PCDHGA9 and CXCR4 unaffected. f A schematic diagram illustrating the regulatory network involving miR-1269a, PCDHGA9, HOXA1, CXCR4, and β-catenin
miR-1269a and PCDHGA9 affect the invasion and metastasis of CRCTo investigate the combined impact of miR-1269a and PCDHGA9 on invasion and metastasis of CRC in vivo, we cotransfected two lentiviruses—Vpcdhga9 or OEpcdhga9 and VmiR-1269a or OEmiR-1269a—into HCT116 cells. Stable cotransfected cells were screened using puromycin and blasticidin. Subsequently, four groups of cells were used to establish lung metastasis models in nude mice. Whole-body fluorescence images revealed that the miR-1269a-only group, devoid of PCDHGA9, exhibited larger and more focal areas of metastatic shadows than the other groups, indicating that the miR-1269a-only group displayed increased invasive and metastatic abilities. Conversely, the group coexpressing miR-1269a and PCDHGA9 demonstrated reduced invasive and metastatic abilities compared with the miR-1269a-only group (Fig. 8a). However, no metastatic shadows were observed in the PCDHGA9-only group (Fig. 8a). Subsequent sacrifice of the mice for lung nodule assessment confirmed that the miR-1269a-only group had more nodules, while PCDHGA9 attenuated miR-1269a’s impact on nodule formation (Fig. 8a, Supplementary Fig. S3c). Similar results were observed in these nodules following immunohistochemical staining for E-cadherin and vimentin (Fig. 8b).
Fig. 8
miR-1269a and PCDHGA9 together affect the invasion and metastasis of CRC in vivo. a Whole-body fluorescence images showing lung metastases across varying conditions of miR-1269a and PCDHGA9 expression. Subsequent photos depicted lung metastases following the sacrifice of these mice. b Immunohistochemical staining of lung metastases was carried out for E-cadherin and vimentin. The immunohistochemical staining highlighted alterations in E-cadherin and vimentin expression in response to different miR-1269a and PCDHGA9 expression conditions. miR-1269a reduced the expression of E-cadherin, but this result was reversed in the condition of high expression of PCDHGA9. Vimentin showed the opposite of E-cadherin
Moreover, we used clinical specimens to establish a combined molecular prognostic model involving miR-1269a and PCDHGA9 and evaluated the expression of miR-1269a in 63 colorectal cancer patient tissues (Supplementary Fig. S3d). However, Spearman’s correlation method revealed no significant correlation between the expressions of miR-1269a and PCDHGA9 in CRC tissues (Supplementary Fig. S3e).
留言 (0)