
記住我


The size, polydispersity index (PDI) and charge of all vesicles were measured and recorded prior to all individual experiments. NISV had a mean size of 1406 ± 22 nm, and a typical PDI of 0.371 and a zeta potential of -29.5mv.
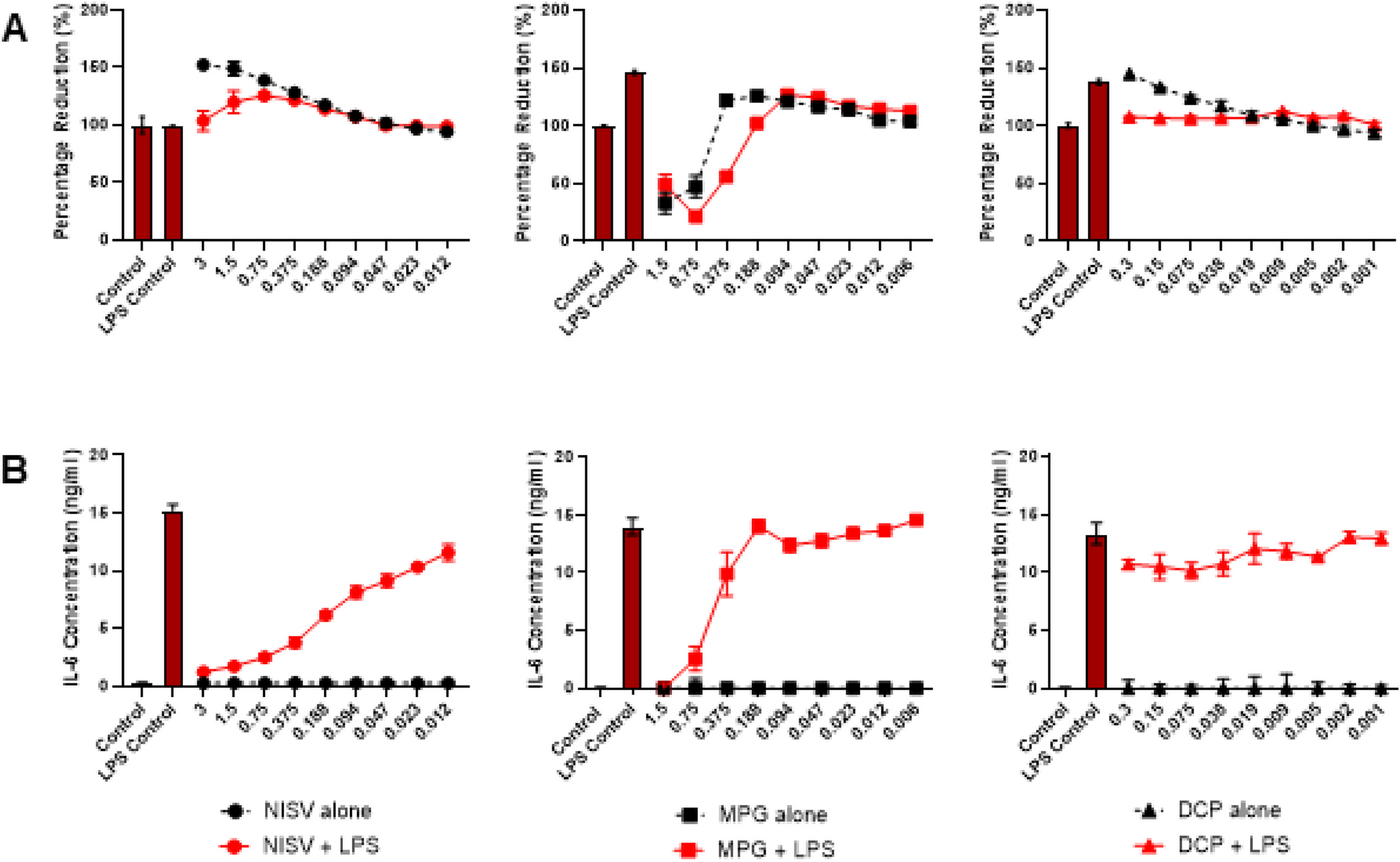
NISV inhibit LPS-induced IL-6 production at non-toxic concentrationsTo measure the anti-inflammatory effects of NISV and ensure minimal toxicity an IL-6 ELISA was performed in tandem with an alamarBlue cell viability assay. This assay measures the reduction of resazurin by the cells and is a proxy for general cell metabolism [20]. The results of the alamarBlue assay on Fig. 1A demonstrates NISV at any concentration used have no negative effects on cell viability. At the higher NISV concentrations used the percentage reduction of alamarBlue by BMDMs was increased suggesting that vesicles augmented cellular metabolism.
Fig. 1
NISV are non-toxic to BMDM and reduce LPS stimulated IL-6 production. 100,000 BMDM were plated per well in triplicate and stimulated with LPS at 3 µg/ml or media, in the controls, followed by treatment with NISV, MPG or DCP at descending concentrations with the controls untreated. A shows alamarBlue reduction by BMDM under these conditions and B shows IL-6 levels in the supernatants 24 h post-treatment. Statistical analysis was performed on prism 8 using two-way ANOVA and Tukey’s multiple comparisons test. * Indicates statistical significance (p < 0.05)
Figure 1B shows the results of the IL-6 ELISA performed with cell supernatants. Minimal IL-6 was produced by unstimulated cells with or without NISV treatment. The LPS stimulated cells induced > 15 ng/ml IL-6 from BMDM. Treatment of LPS stimulated cells with NISV significantly reduced IL-6 production and followed a dose-dependent pattern. To determine if the immunomodulatory effects of vesicles is dependent on their composition, the individual components of NISV were tested to examine their ability to alter IL-6 production.
Each component was ‘prepared’ using the melt method to more closely mimic the NISV preparation. Figure 1A demonstrates that high concentrations of MPG (0.375-1.5 mM) resulted in a reduction of cell viability. This is likely due to the toxicity of surfactant to cells. Concentrations lower than this were non-toxic to cells and at these non-toxic concentrations there were no obvious changes to IL-6 levels. As expected, IL-6 production was reduced where cells were incubated with toxic concentrations of MPG. Cell viability (Fig. 1A) was not profoundly affected by DCP in the presence or absence of LPS. DCP did not notably affect IL-6 levels in LPS-stimulated macrophages. Cholesterol could not be tested in this manner due to its low solubility and the toxicity of solvents required for solubilisation at required comparable concentrations.
NISV have minimal impact on macrophage metabolismThe effects of NISV on glycolysis are shown in Fig. 2A. A number of metabolites and transcripts related to glycolysis were significantly affected by treatment with NISV. Levels of the metabolite D-glyceraldehyde 3-phosphate were significantly (p < 0.05) upregulated in LPS stimulated BMDM consistent with the ability of LPS to induce the Warburg effect. NISV had little effect on D-glyceraldehyde 3-phosphate levels administered to BMDMs, but ameliorated LPS-induced transcript levels reducing relative intensity from 520853 to 197948. NISV increased pyruvate levels in unstimulated cells (relative intensity 930236 to 1502884) and LPS-stimulated cells (relative intensity 669416 to 1056341). NISV increased lactate levels in LPS-stimulated cells (relative intensity 34926857 to 41934274) but had not affect on unstimulated cells.
Fig. 2
NISV alter the metabolism of BMDM following LPS stimulation. 100,000 BMDM were plated per well as 10 replicates and stimulated with LPS at 3 µg/ml or media, in controls, followed by treatment with NISV. Samples were pooled to make 1,000,000 cells/sample. Cells then underwent metabolite extraction and were sent for metabolomics analysis. Pathway diagrams indicated metabolite detection in green and non-detection in black. A shows NISV effects on glycolysis, B shows their effects on the TCA cycle. Statistical analysis was performed on prism 8 using two-way ANOVA and Tukey’s multiple comparisons test. * Indicates statistical significance (p < 0.05)
Enzyme transcripts were also affected by NISV treatment. NISV increased transcription of Aldoart1 in both stimulated (FKPMs of 775 to 1110), and unstimulated cells (FKPMs 966 to 1487). Aldoa transcription was increased in LPS-stimulated cells by NISV (FKPMs from 0.13 to 255). Triosephosphate isomerase, Tpi1, was increased by NISV in both stimulated (from FKPMs 381 to 1189), and unstimulated cells (from 631 to 1186). Pgam1 shows this same phenotype with NISV increasing transcription, from FKPMs of 298 and 235 to 432 and 385 in unstimulated and stimulated cells respectively. Overall, these data indicate that NISV do not have simplistic effect on glycolysis or indeed LPS-induced Warburg effect.
Figure 2B shows the detected metabolites and transcripts associated with the TCA cycle and the alternate M1 macrophage TCA cycle. NISV reduced succinate and itaconate metabolites significantly in LPS-stimulated cells, reducing the relative intensity of succinate from 146453 to 85161 and itaconate from 7017459 to 3712409. In unstimulated cells NISV increased itaconate levels from relative intensities of 139779 to 2670894. Transcription of citrate synthase, Cs, was significantly upregulated by NISV in unstimulated cells, from an FKPM of 55 to 60. Fh1 transcription was reduced in NISV treated unstimulated cells (FKPM of 44 to 33), but not affected in LPS-stimulated cells. Malate dehydrogenase, Mhd2, transcripts were increased by NISV in unstimulated cells (from FKPM from 72 to 78). Overall, these data indicate that NISV has a limited amelioratory effect on LPS-induced remodelling of the TCA cycle.
NISV have significant effects on LPS-induced inflammatory transcriptsTo further elucidate a potential mechanism of NISV anti-inflammatory effects in LPS-stimulated BMDMs, transcriptomic analysis was performed. Cells were first tested for the characteristic reduction in IL-6 production by NISV in LPS-stimulated cells by ELISA, as well as alamarBlue to confirm no toxicity was detected. PCA analysis demonstrated clear separation between all groups tested and consistency within groups (Supplemental Fig. 1). Approximately 22,000 unique transcripts were detected in each sample. LPS stimulation of BMDM cells significantly altered 11,842 transcripts many of which could be easily identified as immunologically important. In comparison, in NISV stimulated BMDM, 3,830 transcripts were significantly altered (Supplemental Table 1).
NISV were found to significantly alter approximately 626 out of approximately 22,000 transcripts detected in LPS-stimulated cells. NISV treatment of LPS stimulated cells was found to most dramatically affect those transcripts associated by ‘Defence Response’ as determined using the GOrilla analysis tool. Included in this group were chemokines, interleukins, tumour necrosis factor α (TNF-α) and their related genes. It was found that NISV significantly downregulates many genes in these respective groups, including genes of important inflammatory effectors; CCL3, CCL4, CCL22, CCL24, CXCL1, CXCL2, CXCL3, CXCL9, CXCL13, IL-1α & β, IL-6, IL-10, IL-12α & β. In addition, NISV were found to significantly downregulate LPS-induced genes of this family, including Tnf (TNF-α), TNF induced protein expression of Tnfaip2 and Tnfaip3 and superfamily members Tnfsf4, Tnfsf9 and Tnfsf15 (Fig. 3).
Fig. 3
NISV treatment downregulates transcripts induced by LPS stimulation. Transcriptomic analysis was performed on RNA extracted from BMDM stimulated with LPS and treated with 1.5 mM NISV, and BMDM controls untreated with NISV, and unstimulated with LPS. A Shows the effects of NISV on the genes responsible for chemokine, interleukin and TNF production in LPS-stimulated BMDM. B shows gene expression involved in LPS signal transduction by the TLR4 signalling cascade as they are altered by NISV treatment in LPS-stimulated cells. Heat maps were made using Prism 8. * Indicates significant change. (p < 0.05)
As NISV were found to regulate a number of transcripts known to be important in the TLR4 receptor signalling cascade, levels of transcripts involved in affected signalling pathways were examined (Fig. 3A). NISV were found to significantly downregulate Cd14 and Map3k7 (TAK1) and significantly upregulate Ticam1 (TRIF), following LPS stimulation. In signal transduction we see no significant effects in the Map3 kinase cascade, the IKK complex or TRAF-3 signalling. Significant changes are shown in transcription factors expression, NF-κB and AP-1, we show significant downregulation of NF-κB subunits 1, 2 and Rel. AP-1 subunits Jun and Fosl2 are significantly altered by NISV treatment, with Jun being upregulated and Fosl2 significantly downregulated. Many of the genes significantly down-regulated by NISV treatment, in LPS-stimulated BMDMs have their expressions controlled by NF-κB, which is activated upon LPS interaction with TLR4. These findings could indicate that NISV exert their anti-inflammatory effects by down-regulating NF-κB (Fig. 3B).
NISV inhibit both Trif and MyD88 dependent signalling pathwaysAs transcript levels for both myD88 and TRIF were affected by NISV, we investigated the effects of NISV treatment on cells stimulated with Pam3csk4 and Poly(I:C) which initiate signalling through myD88 dependent and TRIFF dependent pathways respectively. NISV inhibited IL-6 production by macrophages stimulated with either of these ligands in dose dependent fashions (Fig. 5). Importantly, none of the concentrations of NISV used had a negative effect on BMDM cells as determined by alamarBlue assay (Fig. 4).
Fig. 4
NISV reduce IL-6 response following Poly(IC) or Pam3csk4 stimulation of BMDM. 100,000 BMDM were plated per well in triplicate and stimulated with poly(I:C) at 10 µg/ml, pam3csk4 at 320 ng/ml or media, in the controls, followed by treatment with NISV at descending concentrations with the controls untreated. A shows alamarBlue reduction by BMDM under these conditions and B shows IL-6 levels in the supernatants 24 h post-treatment
NISV inhibits inflammatory cytokine production induced by MyD88 and TRIF dependent signallingA cytometric bead array was used to investigate NISV effects LPS, Poly(I:C) or Pam3csk induced inflammation and to validate the transcriptomic findings for LPS stimulation. Initial analysis found that each of the TLR stimulants induced several inflammatory mediators by the BMDMs. In addition, NISV alone increased certain mediators to a lesser extent (Fig. 5). However, the anti-inflammatory effects of NISV were evident to varying extents irrespective of TLR stimulant. Thus, NISV were found to significantly decrease (p < 0.05) CCL22, G-CSF, IL-6, IL-12p40 and IL-12p70 in LPS-stimulated cells (Fig. 6). They reduced IL-6 and IL-12p40 in Poly(I:C)-stimulated cells and NISV significantly reduced IL-10 and IL-12p40 in Pam3csk4-stimulated BMDM. In all stimulations NISV induced a significant increase in IL-1β. In Poly(I:C) stimulation NISV also increased TGF-β1, and in Pam3csk4-stimulated cells IL-23 and TNF-α production were increased. These data demonstrate that the immunomodulatory abilities of NISV on BMDM are robust, but exhibit subtle, distinct differences depending on the PAMPS used and logically the TLR engaged in macrophages stimulation.
Fig. 5
NISV downregulate inflammatory cytokines from BMDM following LPS stimulation. 100,000 BMDM were plated per well in triplicate and stimulated with NISV, LPS, poly(I:C) or pam3csk4 incubated for 24 h. Supernatants were taken immunological markers measured by LEGENDplex™ cytometric bead array using an AttuneNxT flow cytometer. Heat maps were generated showing Log2(fold change) of vesicle and PAMP treatments compared to an unstimulated control. Statistical analysis was performed using Prism 8 where a two-way ANOVA with Tukey’s multiple comparisons
Fig. 6
NISV alter inflammatory cytokines from BMDM following LPS, Poly(I;C), or Pam3csk4 stimulation. 100,000 BMDM were plated per well in triplicate and were stimulated using 3 µg/ml LPS, 10 µg/ml poly(I:C) and 320 ng/ml pam3csk4, these were then treated with 1.5 mM NISV. After 24 h supernatants were taken and analysed using a cytometric bead array. Heat map shows the Log2(fold change) of NISVcompared to their respective PAMP control. Statistical analysis was carried out using Prism 8 where a two-way ANOVA and Tukey’s multiple comparisons test was performed
留言 (0)