
記住我
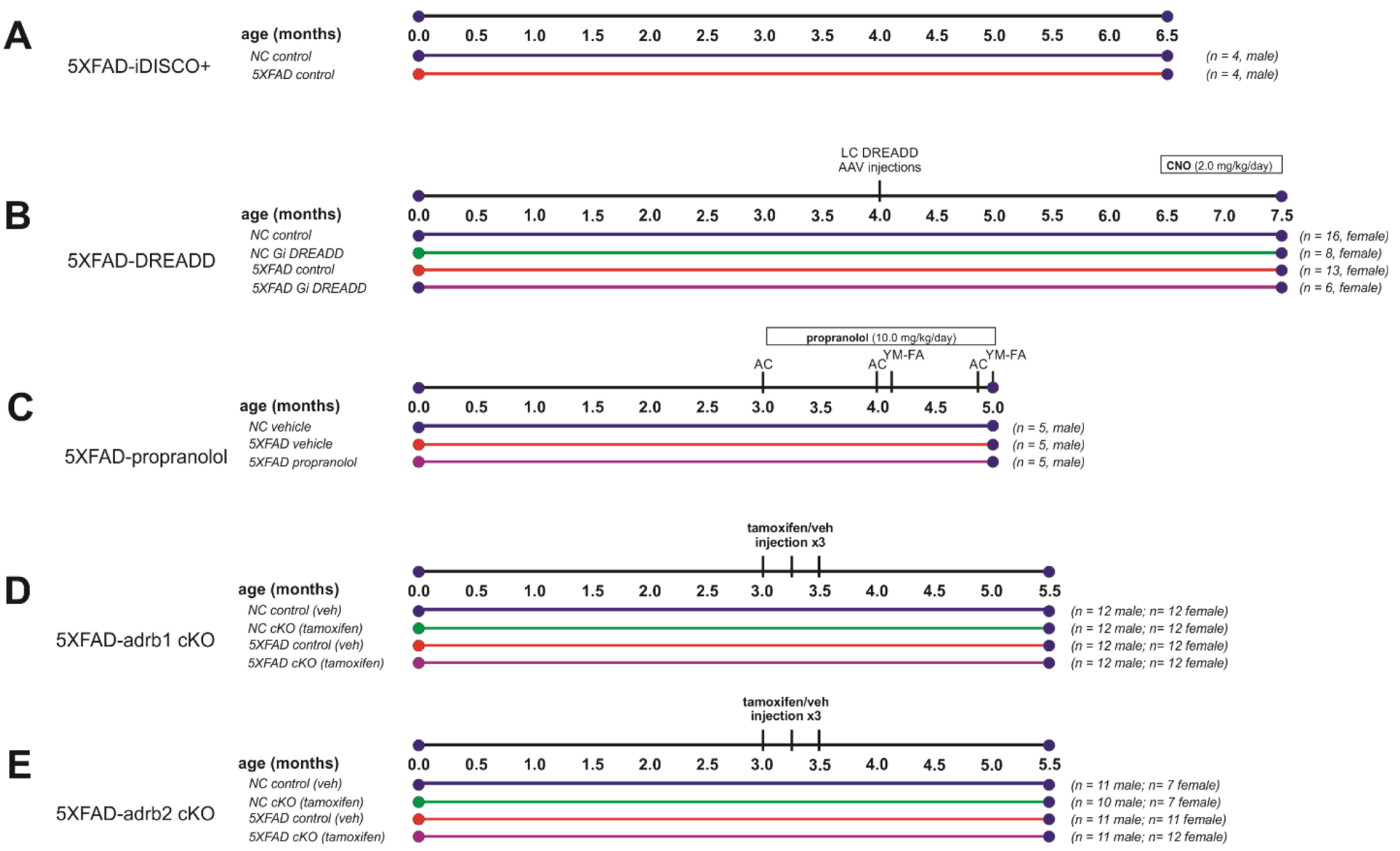

The meninges form a crucial barrier and interface between the CNS and the peripheral environment [21]. This protective layer shields the CNS from physical trauma and pathogen infiltration, situated between the brain and the surrounding bony structures, including the skull and facial bones [30]. The meninges comprise three distinct layers: the outermost dura mater, the middle arachnoid mater, and the innermost pia mater. The arachnoid and pia mater are collectively known as the leptomeninges. In addition to the meninges, the CNS parenchyma is protected by several other barriers, including the blood-brain barrier (BBB), the blood-meningeal barrier (BMB), and the blood-cerebrospinal fluid (CSF) barrier [31, 32]. These barriers typically consist of a single layer of cells connected by tight junctions (TJs), which restrict the free movement of molecules and cells. This selective permeability renders these barriers impermeable to immune cells and most large circulating molecules.
These barrier systems effectively divide the CNS into distinct regions, each with varying levels of accessibility for immune cells and signaling molecules, creating the brain’s unique immune landscape. The main compartments include: (1) Cranial bone marrow; (2) Dural meninges; (3) Leptomeninges; (4) Parenchymal perivascular space (Virchow-Robin space); (5) Choroid plexus; (6) Brain ventricles [30, 33, 34] (Fig. 1A). In this review, we will focus on the meninges and the cranial bone marrow, providing an in-depth analysis of their anatomical structures and functions. This examination will shed light on the complex interplay between these structures and their role in maintaining CNS homeostasis and immune regulation.
Fig. 1
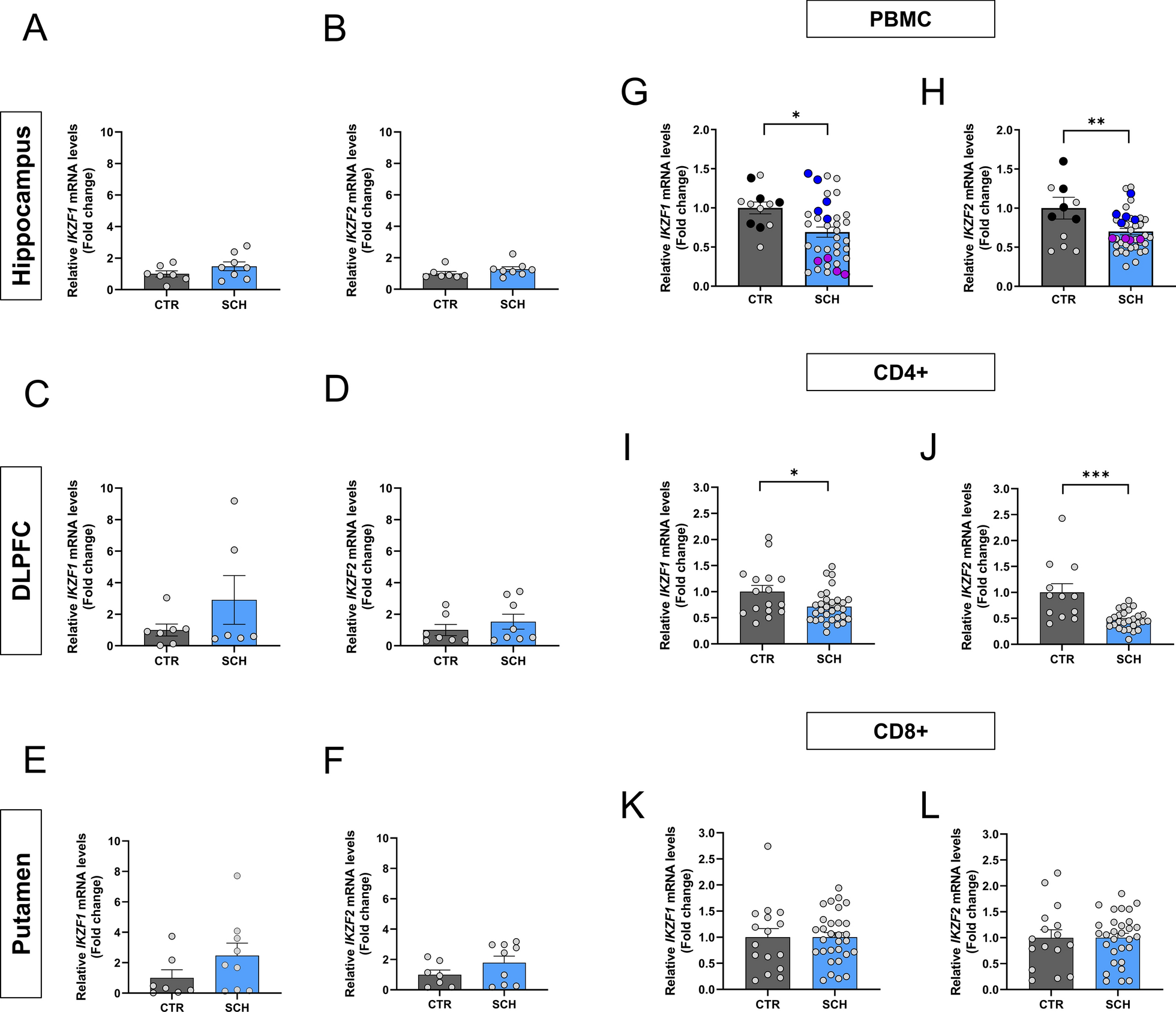
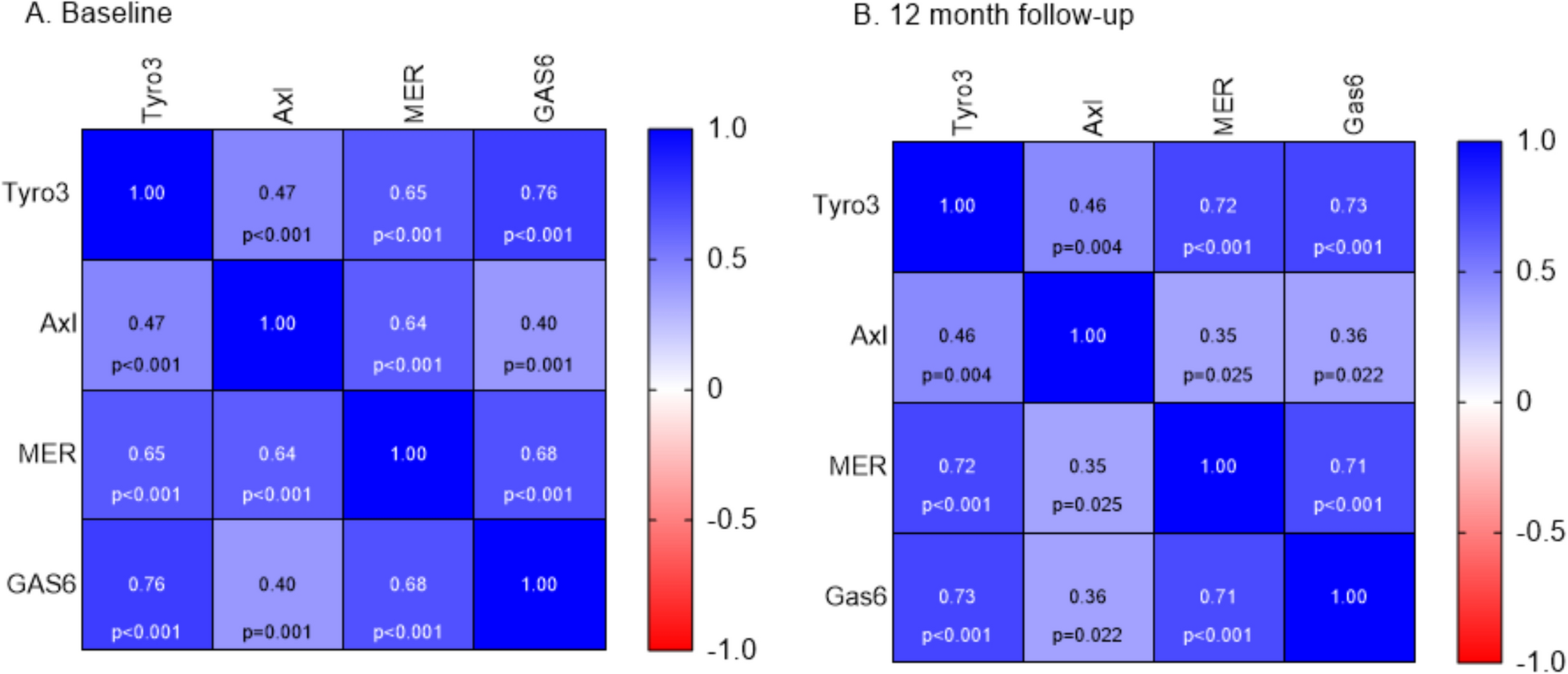
Structure and immune function of meninges. A The meninges, located between the skull and the brain, are composed of three layers: the dura mater, the arachnoid mater, and the pia mater. The dura mater contains a rich distribution of nerves, blood vessels, and lymphatic vessels. The arachnoid mater is thin and transparent, lacking blood vessels and nerves, and it envelops the subarachnoid space, which is filled with cerebrospinal fluid (CSF). The subarachnoid lymphatic-like membrane (SLYM), situated between the arachnoid mater and the pia mater, is proposed to be a potential fourth meningeal layer. The SLYM divides the subarachnoid space into external and internal compartments. It plays a role in restricting the exchange of most small peptides and proteins between the superficial and deep layers of the subarachnoid space, and is also involved in meningeal immunity and fluid drainage. Numerous immune cells are found within the dura mater. In contrast, the leptomeninges and CSF contain fewer immune cells in both number and variety. B The barrier-like structure of the dura mater vasculature is permeable, allowing bidirectional exchange of blood-borne molecules and immune cells with the peripheral circulation. This feature renders the meninges an exceptional platform for immune surveillance. The dura mater is innervated by a network of peripheral nerve fibers, which project to both vascular and non-vascular targets. These fibers can modulate the contraction of dural blood vessels, thereby regulating their permeability. Signals arising from changes in the meninges, such as mechanical deformation or pH variations, can also be transmitted to the central nervous system. C Cells of the arachnoid mater are tightly joined together by tight junctions, preventing molecules and cells from freely passing from the dura mater into the subarachnoid space. This stringent barrier effectively isolates the dura mater from the rest of the central nervous system. Furthermore, arachnoid barrier cells express an abundance of transporters and efflux pumps, which support the regulatory function of this barrier layer over molecular movement. D Neuro-immune crosstalk: Immune cells within the meninges can influence behavior by secreting cytokines that act on neurons, thereby maintaining and modulating neuronal activity. Additionally, these immune cells are capable of responding to neurotransmitters and neuropeptides released by neurons
DuraThe dura, the outermost layer of the meninges, is a robust and dense membrane that envelops the brain [21, 30]. It comprises two distinct layers: the outer periosteal layer, which adheres tightly to the skull, and the inner meningeal layer, lined with flat cells adjacent to arachnoid layer. The vessels channels that collect venous blood from the brain, known as venous sinuses, pass between these two layers [34, 35]. The dura features prominent folds that extend between the cerebral and cerebellar hemispheres. These include the falx cerebri, which separates the cerebral hemispheres, and the tentorium cerebelli, which lies between the cerebrum and cerebellum. These structures play a crucial role in limiting brain movement within the CSF, thereby providing mechanical protection. Notably, the dura is richly vascularized and contains an intricate lymphatic network. It also possesses extensive innervation, making it a highly responsive and dynamic structure. This combination of vascular, lymphatic, and neural elements underscores the dura’s importance not only as a protective layer but also as an active participant in maintaining brain homeostasis and mediating neuroimmune interactions.
Dural blood vesselsThe meninges, particularly the dura, possess a specialized vascular network crucial for immune surveillance [34]. Intracranial vessels can be divided into four main types: around the skull (pericranial type), within skull bones (diploic type), in the brain’s coverings (meningeal type), and inside the brain parenchyma (cerebral type) [36,37,38]. In primates, the dura and skull are perfused by the anterior, middle, and posterior meningeal arteries, which originate from the external carotid artery.
In rodents, the setup is different: the middle meningeal artery comes from the pterygopalatine or stapedial arteries, which branch off from the internal or common carotid arteries. The dura’s vascular network is characterized by anastomosing vessels, including arteries, veins, and fenestrated capillary beds. The primary arterial supply comes from the middle meningeal artery, which nourishes the vascular network between the periosteal dura and the skull.
A distinctive feature of dural blood vessels is the presence of endothelial cells that exhibit a lower expression of TJs, particularly Claudin-5 and occludin [31, 39]. This results in a fenestrated pattern that allows for rapid exchange of large molecules between blood and surrounding tissues, as demonstrated by the quick leakage of dextran and horseradish peroxidase from dural vessels [40, 41]. This increased permeability allows for easier movement of cells and molecules between the meninges and the bloodstream (Fig. 1B). The dura also contains large venous structures, known as dural venous sinuses, which drain blood from both the dura and the brain. The endothelial cells lining these sinuses highly express leukocyte adhesion molecules, including ICAM1 and VCAM1 [16]. The combination of large vascular surface area, high expression of adhesion molecules, low tight junction levels, and relatively slow blood flow [42] creates an environment conducive to cellular and molecular exchange.
These vascular characteristics contribute to the immunological function of the dural vessels, allowing immunoactive soluble factors and cells to enter the dura. Consequently, the steady-state meninges, particularly the dura, harbor a diverse array of immune cells, including macrophages, neutrophils, T cells, B cells, antigen-presenting cells, mast cells, plasma cells, and innate lymphoid cells [16, 17, 43,44,45]. The distribution of these immune cells within the dura is not uniform; they tend to accumulate in specific areas, notably along the walls of the dural venous sinuses. This unique vascular architecture and its associated immune cell populations underscore the dura’s role not just as a protective barrier, but as a dynamic immunological interface between the CNS and the periphery.
Lymphatic vasculature in the duraThe discovery of meningeal lymphatic vessels (mLVs) within the dura in 2015 marked a significant milestone in our understanding of the CNS immunity and fluid dynamics [3, 4]. These vessels express classical lymphatic endothelial cell markers, including VEGFR3, Prox1, Podoplanin, Lyve1, CD31, and CCL21, confirming their lymphatic nature. Meningeal lymphatics form an extensive drainage network that connects the CNS parenchyma to the peripheral immune system. This network comprises dorsal mLVs on the dorsal skull, basal mLVs [5, 46] on the skull base, and an interconnected system within the cavernous sinus [47] (Fig. 2D). These vessels serve crucial functions in draining CSF, antigens, and immune cells, thereby clearing waste and coordinating immune responses in the CNS [48].
Fig. 2
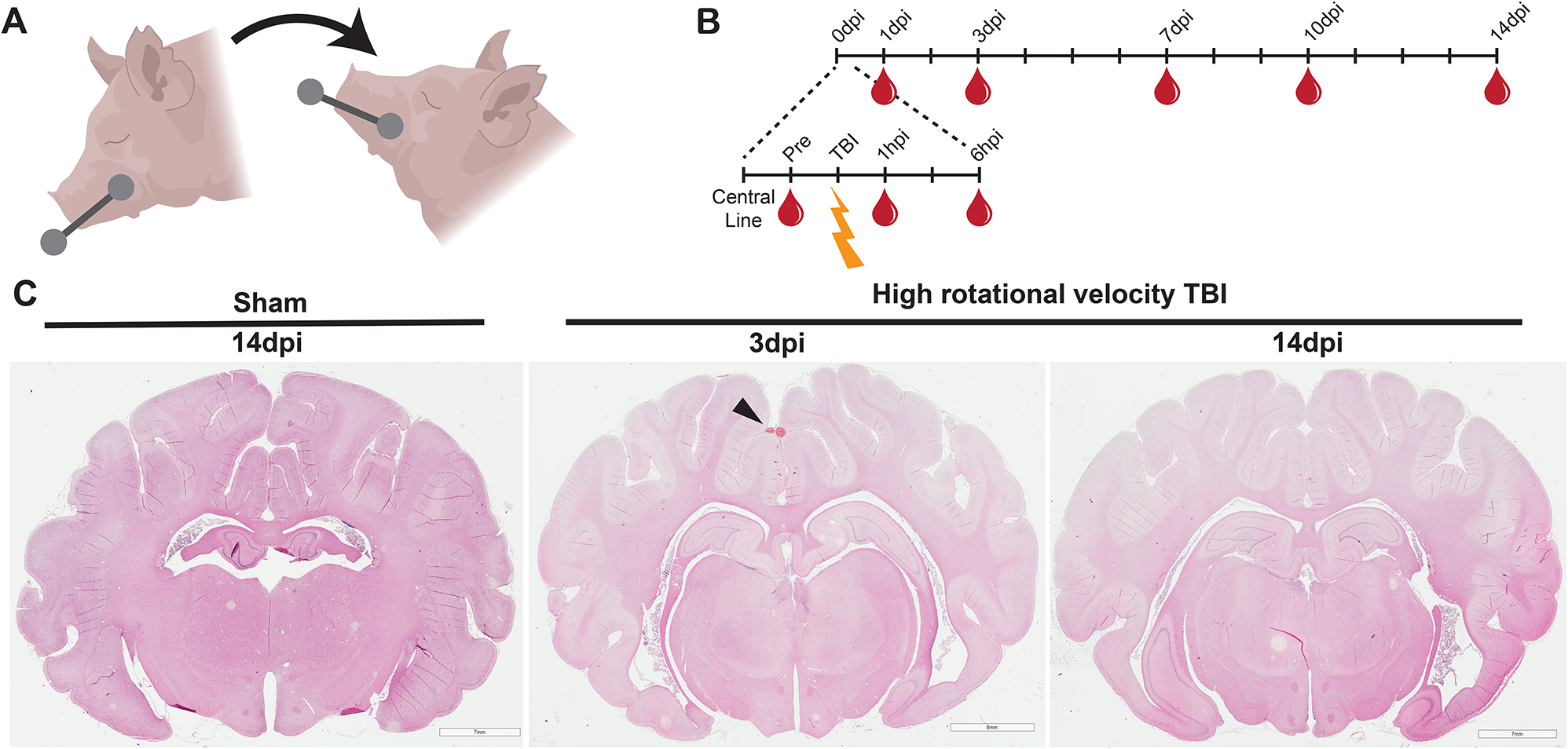
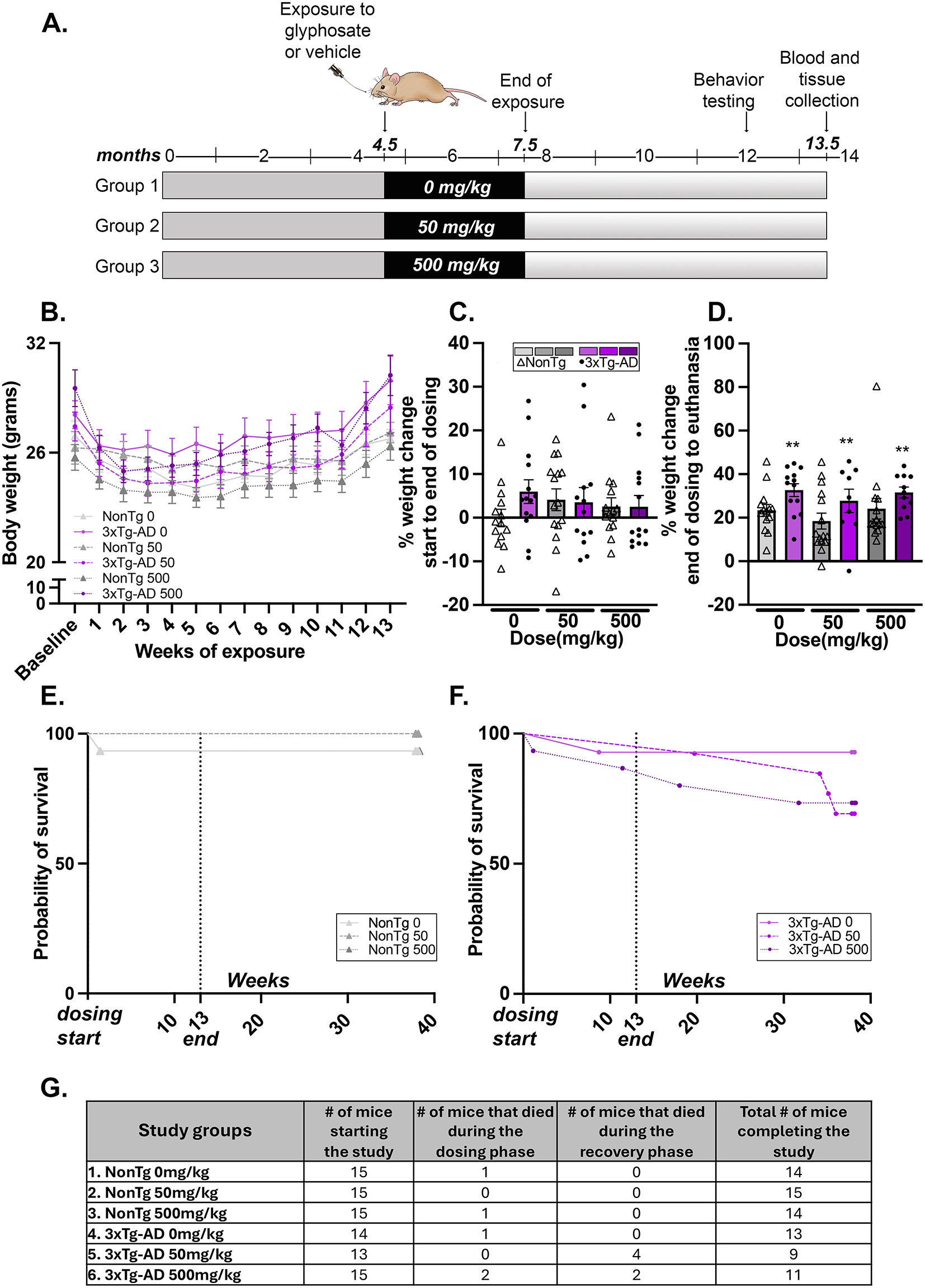
Specialised structures at the meningeal interface. A Dural-Associated Lymphoid Tissues (DALT). DALT represents a structured lymphoid network interwoven with the barrier-like vasculature within the dura mater. The red box highlights the rostral-rhinal hub, which connects with multiple veins and the bone marrow of the skull. B Arachnoid granules. The arachnoid mater forms numerous villous protrusions on both sides of the superior sagittal sinus, which extend into the sinus and are referred to as arachnoid granulations. These structures may be involved in the reabsorption of cerebrospinal fluid (CSF) into the venous blood. Arachnoid granulations can be classified into five types based on their morphology and their relationship with the dura mater, each contributing to varying degrees to CSF flow and meningeal immunity. C Arachnoid cuff exit (ACE) points. As the bridging vein traverses the arachnoid barrier, it creates discontinuities, forming channels that connect the subarachnoid space with the dura mater. A’ illustrates that the bridging vein is located within the arachnoid mater. B’ shows that at the ACE point, the bridging vein is positioned outside the arachnoid mater. D Cranial lymphatic structures. Cerebral lymphatics can be categorized into meningeal lymphatics, basal mLVs, and nasal lymphatics and nasopharyngeal lymphatic plexus. Through these channels, CSF, carrying immune cells, cytokines, and metabolic waste, is drained from the subarachnoid space into the deep cervical lymph nodes (dCLN)
The anatomical distribution of mLVs correlates with their distinct morphological characteristics and functions. Dorsal mLVs, which run parallel to dural venous structures like the superior sagittal (SSS) and transverse sinuses (TS), have small, unbranched lumens without valves and discontinuous endothelial cell connections [3, 4]. In contrast, basal mLVs, located along the parietal sinuses (PSS) and sigmoid sinuses (SS), feature larger, branched lumens with valves and oak leaf-shaped endothelial cells [5, 46]. Both the dorsal mLV and the basal mLV can drain CSF out of the brain. However, the basal mLV is morphologically superior and is located adjacent to the subarachnoid space, so it is considered to be more effective in draining large molecules from CSF. Together with dural venous sinuses, these lymphatic vessels form dural immune centers, facilitating CSF sampling, antigen uptake, and presentation to T cells [16]. This arrangement plays a crucial role in establishing the neuroimmune interface. Additionally, the potential interconnection between meningeal lymphatics and the lymphatic systems of the nasal and nasopharyngeal regions suggests a continuous drainage and immune defense network extending from the CNS to the upper respiratory tract.
The mLVs network has been implicated in various neurological conditions, including aging [49], Alzheimer’s disease [50, 51], Parkinson’s disease [52], traumatic brain injury [53], subarachnoid hemorrhage [54], CNS viral infections [55], and brain tumors [56]. Changes in mLVs transport capacity can significantly influence disease progression. For instance, the deterioration of mLVs function with age or in Alzheimer’s disease may contribute to cognitive impairment and exacerbate neurological damage. Moreover, the extensive distribution of mLVs within the dura offers promising avenues for transcranial nerve modulation therapies. Treatments targeting the lymphatic system show great potential for managing neurological conditions. Enhancing lymphatic drainage to clear toxic molecules from the CNS could improve cognitive function in age-related disorders. Future directions in neurological disease treatment may include developing non-invasive modalities to regulate mLVs function, such as pharmacological interventions and photostimulation techniques [51, 57, 58]. This evolving understanding of mLVs has fundamentally altered our perspective on intracranial fluid dynamics and brain immunity, opening new avenues for research and potential therapeutic strategies in neurological disorders.
Dural-associated lymphoid tissues (DALT)Recent research by Fitzpatrick et al. has revealed a network of organized lymphoid structures within the dura of mice and humans, termed dural-associated lymphoid tissues (DALT) [59] (Fig. 2A). This groundbreaking study employed advanced techniques, including micro-CT imaging, single-cell RNA sequencing, and B cell receptor (BCR) sequencing, to provide a comprehensive description of these structures. DALT is intricately interwoven with the fenestrated vasculature of the dura, with its most complex components located in the rostral-rhinal and basal olfactory venous hubs. The rostral-rhinal DALT, also known as the rostral-rhinal venolymphatic hub, is directly connected to cranial marrow vessels via diploic veins. It also receives vascular input from multiple sources, including the rostral-rhinal sinuses, olfactory sinus, superior sagittal sinus (SSS), bridging veins, and cortical veins. These venolymphatic hubs host diverse immune cell populations, including various B cell subsets, plasma cells, T follicular helper (TFH) cells, and T follicular regulatory (TFR) cells. During pathogen invasion, interactions between germinal center B cells and TFH cells, mediated by IL21-IL21R and CD28-CD86 pathways, are intensified. B cells activated within DALT subsequently migrate to the sinus wall, bolstering local immunity in the venous sinus.
This discovery presents a novel model for understanding CNS immunosurveillance, highlighting DALT’s strategic anatomical position for acquiring both local and whole-brain antigens. DALT can harvest antigens from blood and nasal passages to generate localized immune responses, supporting the production of class-regulated, high-affinity antibodies in response to viral challenges. These venous lymphoid hubs facilitate rapid expansion of antigen-specific immune responses, potentially offering defense against foreign pathogens threatening the CNS parenchyma. The role of DALT in supporting humoral immunity within the meninges challenges the traditional view of the meninges as a simple barrier. Instead, it reveals a complex, immunologically active interface between the CNS and the periphery. This paradigm shift in our understanding of meningeal structures opens new avenues for research into CNS immunity and potential therapeutic approaches for neurological disorders.
Dural nerveThe meninges, particularly the dura, are characterized by extensive neural innervation [60, 61]. This innervation network comprises peripheral sensory and autonomic nerves that target both vascular and non-vascular structures. The trigeminal nerve, primarily its first branch, is the main supplier to the dura, with additional contributions from its other branches. Notably, the tentorial nerve, a branch of the first division, innervates the upper dura mater, the superior sagittal sinus, the transverse sinus, and the parietal branches of the middle meningeal artery.
Trigeminal ganglion projections extend to the middle cerebral artery (MCA) and form intricate loops with the middle meningeal artery (MMA) [62]. These neural fibers co-innervate various meningeal blood vessels and resident cells (Fig. 1B). Interestingly, some trigeminal nerve afferents form loops that terminate in the cranial bone marrow or traverse the skull to end in the external periosteal layer, potentially facilitating communication across the meningeal barrier.
The meninges constitute a highly sensitive sensory system. Sensory fibers within the meninges respond to changes in temperature, pH, and mechanical pressure, providing critical feedback about meningeal status [55, 63]. These afferents serve as key sensory transducers in proprioception and vestibular sensation, capable of detecting physiological meningeal deformation and possibly changes in intracranial pressure. This sensitivity may play a protective role during head impacts, with abnormal meningeal stretching potentially signaling to prevent injury during rapid head movements.
The meningeal nerve supply is composed of thousands of sympathetic, parasympathetic, and sensory fibers, some of which are myelinated. This network forms a localized chemical defense system. Trigeminal nerve projections to the dura and pons contain vasoactive neuropeptides such as calcitonin gene-related peptide (CGRP), substance P, and pituitary adenylate cyclase-activating polypeptide (PACAP) [55, 64]. When activated, these neuropeptides are released to detect and limit tissue damage by modulating vascular function and signaling the brain. Dural blood vessels exhibit heightened sensitivity to mechanical and chemical stimuli compared to deeper brain vessels. Their responsiveness is complex: increased luminal pressure, neurotransmitters like norepinephrine, and neuropeptides such as neuropeptide Y can induce vasoconstriction. Conversely, electrical stimulation, certain neuropeptides (including CGRP and substance P), acetylcholine, histamine, and serotonin can promote vasodilation.
There are numerous immune cells and neural cells within the meninges, and their direct physical contact offers intriguing new research avenues for the concept of neuroimmune (Fig. 1D). Under normal circumstances, macrophages in the meninges detect, attack, and recruit other immune cells to combat bacterial invasion. The study by Chiu et al. highlights a critical neuroimmune axis in the meninges where bacteria exploit neurochemical signaling to suppress immune responses [65]. Specifically, bacteria like Streptococcus pneumoniae release toxins that activate pain neurons within the meninges. The Nav1.8 + nociceptors are activated, leading to the release of the signaling molecule CGRP by these pain neurons. CGRP downregulates the expression of chemokines by meningeal macrophages through receptor activity-modifying protein 1 (RAMP1), inhibiting neutrophil recruitment and thereby suppressing the host’s immune defenses, exacerbating bacterial meningitis. This intricate interplay between neural and immune components underscores the complex regulatory mechanisms at work in the meningeal microenvironment. Furthermore, the activity of resident mast cells in the meninges is tightly regulated by neurotransmitters and neuropeptides to maintain central nervous system homeostasis [66].
ArachnoidThe arachnoid mater, the middle layer of the three meningeal layers, is a complex structure composed primarily of avascular connective tissue. It consists of multiple cell layers, with an inner layer 1 to 4 cells thick [18, 67]. A distinctive feature of the arachnoid mater is its trabeculae, formed by fibroblast-like cells [15, 65], which span the subarachnoid space and extend to the pia mater. The subarachnoid space (SAS), a CSF-filled cavity between arachnoid and pia, serves critical functions. It houses large arteries that penetrate the brain parenchyma and provides a cushioning effect, protecting the brain from impact injuries. Moreover, CSF within SAS is crucial for brain buoyancy, effectively reducing the brain’s weight and preventing compression under its own mass.
Compared to the dura, a key characteristic of the arachnoid mater is its relative impermeability. This property stems from its unique cellular composition and junctional complexes. In mice, arachnoid barrier (AB) cells originate from mesenchymal precursor cells but begin expressing E-cadherin and Claudin 11 around embryonic day 14 (E14) [15, 68]. The arachnoid mater forms an extensive barrier through tight junctions (TJs) containing Claudin-11 and adherens junctions (AJs) containing E-cadherin [68,69,70]. This barrier effectively separates and regulates the dura and subarachnoid space, restricting bidirectional molecular movement between them (Fig. 1C). For instance, 10 kDa dextran and chemokines (8–10 kDa) are blocked by the arachnoid mater, preventing their movement from the dura to the SAS.
While the arachnoid’s impermeability is fundamental in safeguarding the CNS, it is important to acknowledge that it also demonstrates a degree of selective permeability, particularly in the context of its cellular transport mechanisms. The presence of various transporters and efflux pumps in arachnoid barrier cells enhances their regulatory function. These include ATP-binding Cassette (ABC) Transporters such as ABCB1 (P-glycoprotein), ABCC4 (MRP4), and ABCG2 (breast cancer resistance protein), as well as slc22a6 and slc22a8 [71,72,73,74,75,76,77]. This array of transporters likely contributes to the selective permeability of the arachnoid barrier, allowing for finely tuned regulation of substance exchange between the blood and CSF. This complex structure and function of the arachnoid mater underscore its critical role in maintaining the unique environment of the central nervous system, balancing protection with selective permeability.
Arachnoid granulesWhile the arachnoid barrier creates a partition between the CNS and the dura, communication between these compartments is essential for toxin molecule clearance and immune surveillance. In humans and other large mammals, this communication occurs through arachnoid granules (AGs), which are specialized protrusions of the arachnoid mater. Arachnoid granules, also known as arachnoid villi or Pacchionian granulations, were first described by the Italian anatomist Antonio Pacchioni [78]. These structures have traditionally been described as penetrating the dura mater and facilitating CSF drainage into the dural venous system, but this understanding continues to evolve, reflecting a broader range of functions [79, 80]. Interestingly, AGs do not appear in smaller mammals like rodents, implying significant differences in fluid dynamics and immune interactions between species. This absence may suggest different pathways for CSF clearance and immune surveillance in these species.
Recent research by Shah et al. has provided detailed structural and cellular analyses of AGs, expanding our understanding of their potential roles beyond simple CSF drainage [81] (Fig. 2B). This study suggests that AGs may function as lymphatic conduits, connecting the bone marrow to the arachnoid stroma. This connection could play crucial roles in lymphatic-lymphatic coupling, CSF antigen clearance, homeostasis maintenance, and mechanisms underlying neurological diseases. AGs are classified into five types based on their association with the dura and venous sinuses: intrasinus, stromal, transdural, epidural, and subdural. Structurally, AGs consist of an outer capsule and an inner stromal core, encapsulated by a membrane composed of arachnoid cells. These structures exhibit significant heterogeneity in size, shape, location, and internal architecture.
Internally, AGs present as spongy cavities comprising complex lacunae, crypts, and sinuses. These cavities are rich in diverse immune cells, including macrophages, neutrophils, mast cells, dendritic cells, CD4 + and CD8 + T lymphocytes, B lymphocytes, and plasma cells. Notably, stromal AGs embedded in the dural matrix show an enrichment of immune cells, particularly MHCII + antigen-presenting cells and CD4 + T cells. This cellular composition suggests potential antigen-presenting cell interactions within these structures. The architecture of AGs undergoes significant age-related changes, including increased size, enhanced lobulation, and loss of vesicle integrity. These structural alterations may have important implications for AG function over the lifespan.
In essence, AGs serve as sophisticated filtering channels for CSF, cellular debris, and immune cells. This multifaceted role suggests that AGs play a more complex part in brain-immune interactions than previously thought. Their unique structure and cellular composition position them as key players in maintaining CNS homeostasis and potentially in the pathogenesis of neurological disorders. This evolving understanding of AG function opens new avenues for research into CNS-immune system interactions and may provide novel insights into the mechanisms underlying various neurological conditions.
Arachnoid cuff exit (ACE) pointsThe arachnoid mater’s relative impermeability and transport capacity enable it to regulate the exchange of molecules and factors between the CNS and its surroundings. However, the mechanisms by which the arachnoid barrier balances separation and communication have remained largely elusive. Recent research by Leon C. D. Smyth and colleagues has unveiled a unique structure within the arachnoid mater that sheds light on this complex interplay. Blood draining from the CNS must pass through bridging veins to reach the venous sinuses. These veins traverse the subarachnoid space and penetrate the arachnoid barrier, creating disconti
留言 (0)