
記住我
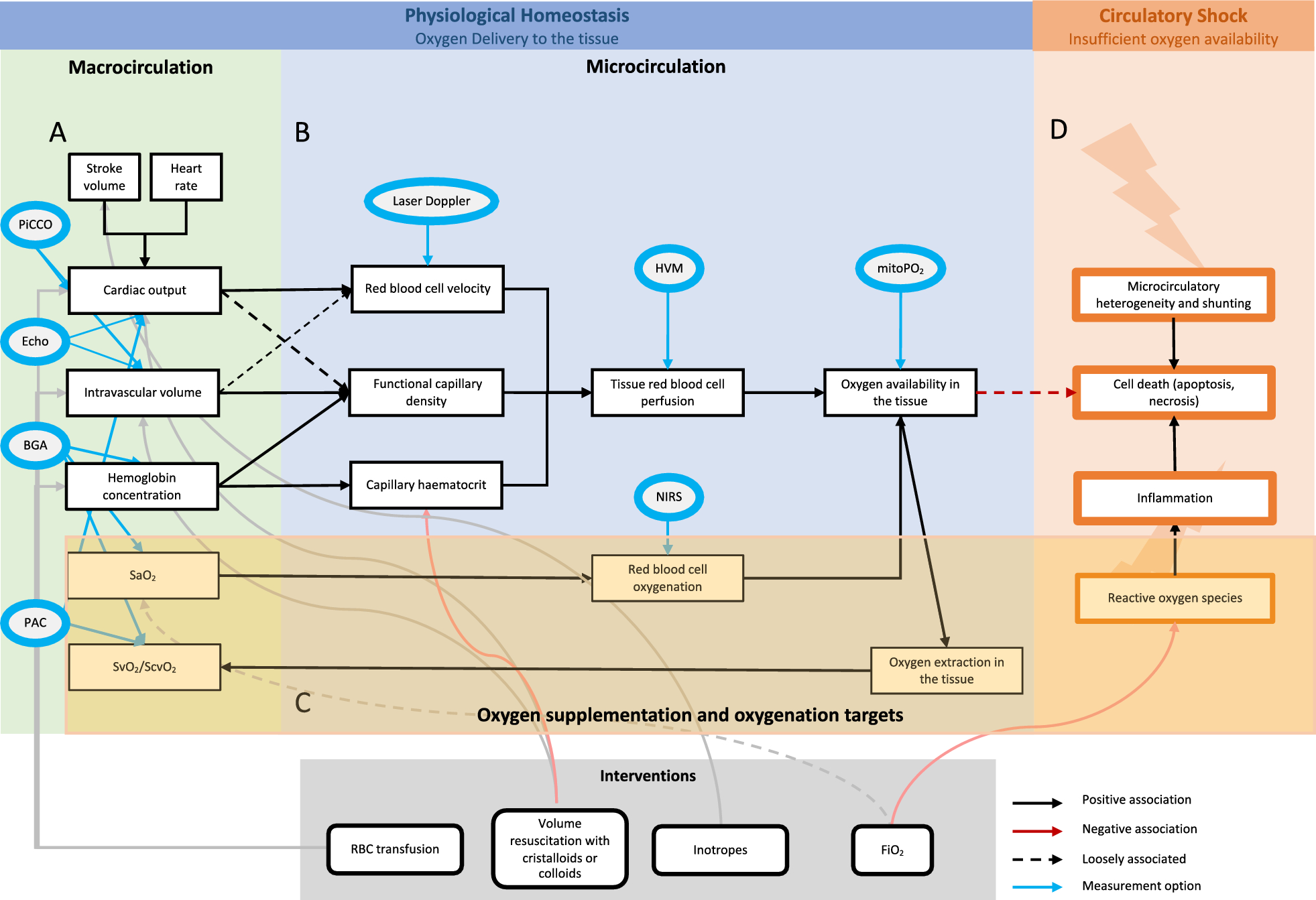


Circulatory shock, which is defined as a life-threatening state of circulatory system failure associated with decreased tissue perfusion, leading to inadequate oxygen delivery (DO2) to meet cellular metabolic demands, remains a common condition with high morbidity and mortality in the intensive care unit (ICU) [1, 2]. Rapid restitution and maintenance of adequate tissue perfusion and oxygenation is the main treatment goal in critically ill patients in shock [3]. The determinants of global DO2 are the cardiac output (CO), the hemoglobin (Hb) concentration and the oxygen saturation in the arterial blood (SaO2). Hemodynamic management in the ICU thus aims to optimize these three physiological variables (Fig. 1A). However, defining targets for each of these variables to rapidly restore tissue perfusion while avoiding adverse effects associated with over-resuscitation (Fig. 1D), remains a challenge.
Fig. 1
Overview of tissue oxygenation. Physiological homeostasis of oxygen delivery to the tissue depends on macrocirculatory (A) and microcirculatory (B) parameters. The macrocirculatory parameters, such as Hb concentration, SaO2, CO and intravascular volume rely on the microcirculatory function. Circulatory shock (D) with insufficient oxygen availability in the tissue is due to reactive oxygen species, inflammation and microcirculatory heterogeneity leading to cell death. The review aims to focus on oxygenation targets, representing a delicate balance between risks and benefits (C). Interventions to influence the different parameters are shown in grey. The measurements options are shown in blue. PiCCO Pulse Contour Cardiac Output, Echo Echocardiography, BGA Blood Gas Analysis, PAC Pulmonary Artery Catheter, HVM Hand-held Vital Microscopy, mitPO2 mitochondrial PO2, NIRS Near-Infrared Spectroscopy
Current resuscitation protocols often emphasize an increase in CO. Interventions are guided by volume or inotrope responsiveness of the stroke volume (SV), a concept based on the Frank-Starling relationship [4, 5] and contractility. They are implemented in various ways in clinical practice [6,7,8]. The focus of the resuscitation is primarily the macrocirculation (Fig. 1A) although in circulatory shock the coherence between the macro- and the microcirculation is often uncoupled. This is shown by an absence of increase in tissue perfusion even though SV might still be responsive to a hemodynamic intervention. Thus, even in presence of persistent fluid responsiveness continued volume resuscitation may be associated with a negative effect on tissue perfusion [9] and with worse outcome [10, 11]. In the case of volume resuscitation, this can be partly explained by a reduction in tissue perfusion with red blood cells through hemodilution related decrease in capillary hematocrit. Without knowledge of the determinants of tissue perfusion and oxygenation, the optimal target for CO remains unknown and may vary from person to person [12]. Additionally, the relationship between DO2 and consumption in sepsis and septic shock has been found to depend on the presence of microcirculatory shunting in addition to mitochondrial dysfunction [13]. Recent technological developments that allow direct bedside assessment of microcirculatory function could open up the possibility of targeting microcirculation [14,15,16] and put individualized, tissue red blood cell perfusion focused therapy within reach [17, 18].
The second determinant of global DO2, Hb concentration, directly facilitates oxygen transport in the blood as oxygen is very poorly soluble in blood plasma (< 3%). Anemia due to various causes is common in critically ill patients. However the transfusion thresholds for these patients are mainly based on two trials, the TRICC [19] and TRISS [20] trial. The trials showed a similar 90-day mortality comparing a Hb of 7 g per deciliter (g/dl) and of 9 g/dl. In patients with septic shock, mortality at 90 days, rates of ischemic events and use of life support were similar in those with a higher Hb target and those assigned to blood transfusion at a lower threshold; the latter group received fewer transfusions [20]. Following these trials Hb targets between 7 and 8 g/dl were defined for most patients, depending on some general additional factors, such as hemodynamic instability, acute bleeding, or risk factors such as previous surgery or coronary artery conditions [21,22,23]. However, these studies do not fully represent the heterogeneous population of critically ill patients suffering from different types of circulatory shock. Nevertheless, the commonly used Hb concentration targets provide little individualization and often do not consider its role in the restoration of tissue perfusion and organ function (e.g., kidney) in patients with circulatory shock [24].
In terms of optimizing the risk–benefit ratio of hemodynamic stabilization of patients with circulatory shock, oxygen supplementation to increase oxygen content per blood volume, in absence of lung disease, may be the most important to consider (Fig. 1C). A stronger focus on the risks associated with the intervention is desirable, because on the one hand, changes in blood oxygenation within the physiological range of oxygen saturation, according to the dissociation curve, only marginally influence global oxygen supply. On the other hand, supramaximal pulmonary and blood oxygenation can be associated with an increased potential for negative effects. However, increasing acidosis due to tissue hypoperfusion may result in increased DO2 due to the Bohr effect on the dissociation curve. Previous studies have demonstrated that critically ill patients often show high SaO2 values even though there are indications that the relationship between SaO2 and mortality likely is U-shaped [25]. The difficulty in defining SaO2 targets may thus represent a risk for hyperoxia based on fear of hypoxia and avoiding hyperoxia could represent a promising strategy to improve patient management.
This narrative review aims to explore the factors influencing decision-making regarding oxygenation targets in critically ill patients with circulatory shock. It examines the risks and benefits of oxygen supplementation by assessing the physiological basis for DO2 and the regulatory mechanisms designed to counteract deficiencies. By providing an overview of the relevant literature, we aim to support decision making at the bedside and provide an outlook on future trends.
Oxygen delivery to the tissues is the basis for all processes of lifeTo reach the current understanding of the role of oxygen in sustaining life has taken many centuries of research. Oxygen is essential for modern metazoan organisms, which emerged around 300 million years ago, coinciding with the significant rise of oxygen levels in Earth’s atmosphere [26]. Oxygen was independently discovered by the English chemist Joseph Priestley and Carl Wilhelm Scheel around 1774 [27], and was named by Antoine Lavoisier in 1778. The “Pneumatic Institution”, founded 1798 in Bristol, was one of the first places where the effects of oxygen on the human organism were examined in the setting of different illnesses. In collaboration with James Watt and Humphry Davy many new methods to deliver oxygen to patients were developed. The research was accelerated at the beginning of the twentieth century with the discovery of oxygen tensions as partial pressure by Adolf Fick and Paul Bert. But it was not until 1917 that John Scott Haldane, following a coal mine explosion, developed the first face mask with a possibility to adjust the administration of oxygen [28]. However, the administration of supplemental oxygen is only the first step, as the oxygen must find its way to the tissue, where oxidative phosphorylation takes place. Oxygen rich blood travels through a network of branching vasculature and is distributed in the tissue by the microcirculation, consisting of arterioles, capillaries, and post-capillary venules with a diameter below 20 µm. The red blood cells, which measure between 3 and 6 µm, travel through the capillaries in a single file fashion and provide oxygen via convection and diffusion [15]. The former occurs through the movement of Hb-bound oxygen molecules from the red blood cells in the capillary network to the mitochondria to fulfil their metabolic function [29]. In this process, the high affinity of the cytochrome c oxidase, the enzyme that reduces oxygen to water, to oxygen plays an important role in maintaining homeostasis by binding oxygen over a wide range of local oxygen pressures in the mitochondria, as low as 0.3–1.0 kPa. This remarkable property forms the basis for the oxygen conformance theory, which states that only at the extremely low end of tissue oxygenation, oxygen demand becomes dependent on supply. In other words, the functionality of oxidative phosphorylation as the basis of all life, can be maintained in the most extreme of conditions [30, 31].
Physiologic adaptation to hypoxemia demonstrates the adaptability of the pulmonary and systemic microcirculationIn line with the importance of maintaining oxidative phosphorylation, the physiological processes along the oxygen supply chain are aimed at avoiding hypoxemia and hypoxia, the former referring to low blood oxygen content, and the latter, to low oxygen levels in the tissue. Genetic and physiological adaptation mechanisms to hypoxia ensure the maintenance of the homeostasis in states of external limitation of oxygen supply, and internal causes of tissue mal perfusion due to systemic disease. However, before understanding the role of hypoxia in disease, isolated models of tissue hypoxia were needed to examine these intrinsic mechanisms. Early research on adaptation to hypoxia was performed by Paul Bert in his compression chamber at the University of Sorbonne in Paris in the nineteenth century. In the following twentieth century subsequent field research was extended to high altitude locations around the world [32]. As partial pressure of oxygen decreases with ascent to high altitudes, the human body relies on an intricate system to detect the lower oxygen availability and react to it to maintain adequate tissue oxygenation. Some of these mechanisms focus on the functioning of the lungs, others on the systemic organs. In general, all animals express hypoxia-inducible factor (HIF) 1, composed of HIF-1α and HIF-1β, and vertebrates also produce HIF-2 and HIF-3. HIF-1 and HIF-2 can activate gene transcription which in turn regulates systemic DO2 and utilization, the role of HIF-3 is less well known. HIF-1 is regulated by oxygen-dependent hydroxylation by the von Hippel-Lindau protein. The O2-dependent binding is inhibited during hypoxic conditions and the HIF-1 activates some and inhibits other genes. At the tissue level, hypoxia leads to angiogenesis via the regulation of vascular endothelial growth factor and to a shift to anaerobic metabolism via the induction of glycolysis and glucose transporters. At the same time HIF-2 regulates several genes that control erythropoiesis [33]. Moreover, HIF are crucial in a multitude of mechanisms protecting cells from oxidative stress by increasing antioxidant production and decreasing oxidant production [34]. While HIF effectively regulates medium- and long-term responses on a cellular level, immediate physiological adaptation is needed to provide acute adaptation to hypoxia.
In order to regulate the function of the cardio-respiratory system during hypoxia, oxygen levels are sensed rapidly at the glomus caroticum, which is located at the bifurcation of the internal and external carotid arteries. The chemoreceptor tissue, which contains type I neuronal glomus cells and type II sustentacular, glia-like cells, is sensory innervated by the carotid sinus nerve. The exact mechanism to detect hypoxia in these cells is not yet found and still under debate. It is assumed that hypoxia depolarizes the glomus cells through a inhibition of K + cannels and that the subsequent calcium-dependent release of excitatory neurotransmitters increases the neuronal activity [35]. In this way, cardiovascular and respiratory responses are triggered and / or modulated. In addition, different parts of the circulatory system have intrinsic regulation mechanisms. The pulmonary circulation responds with vasoconstriction of the pre-alveolar arterioles to a decrease of alveolar oxygen partial pressure. The effect was first described by Bradford and Dean in 1889 and was subsequently named Euler–Liljestrand-reflex [36]. Its rapid onset results from constriction of the small intrapulmonary arteries, mainly the pre-capillary vessels but also, to some extent, the post-capillary venules [37]. The sensory mechanism to detect alveolar hypoxia seems to be within the mitochondria of the smooth muscle cells of the pulmonary arteries [38]. Thanks to this mechanism, a ventilation-perfusion mismatch can be avoided. In global hypoxia, such as at high altitude or with diffuse lung damage, a diffuse Euler-Liljestrand-reflex leads to an increase of pulmonary artery pressure [39]. In the systemic circulation, on the other hand, the focus is to optimize oxygen availability in the tissues (Fig. 1B). Autoregulation of arterial tone plays an important role in the regional distribution of blood flow [40]. An increase in the activity of the sympathetic nervous system during acute hypoxemia, and above all a reduction of the activity of the parasympathetic nervous system in the following weeks, appears to be responsible for an increase in heart rate [41]. Simultaneously changes in plasma volume appear to cause a decrease in SV which ultimately leads to a constant CO [32]. These changes are often confounded by additional factors such as exercise or hypovolemia. Furthermore, systemic vascular tone and systemic vascular hindrance have been found to remain unaffected during ascent to high altitude. Recent observations have led to a deeper understanding of the mechanisms to increase DO2 to the tissue during hypoxic exposure. In a large study of healthy volunteers ascending to 7124 m, recruitment of pre-existing capillaries was identified as the main physiological response to increase microcirculatory oxygen extraction capacity at high altitude [42]. A variability in the response of the microcirculation has been described in different organs [43, 44]. Dark field microscopy images of the sublingual microcirculation recorded in healthy volunteers at sea level and after 2 weeks at 7042 m, representative for the response mechanisms to hypoxia, are shown in Fig. 2A, B.
Fig. 2
Sublingual microcirculation images. Representative images of the sublingual microcirculation before and after the topical application of nitroglycerin, during exposure to extreme altitude, and in critically ill COVID-19 patients. The sublingual microcirculation shows a similarly reaction to hypoxia in healthy volunteers at high altitudes, and critically ill COVID-19 patients. The application of a topical nitroglycerin in healthy volunteers leads to an increase of capillary density that is similar to adaptation to high altitude. Adapted from [42, 45]
Effects and adaptation to hypoxemia in critically ill patientsDifferent to volunteers at high altitude, critically ill patients in circulatory shock often present with insufficient tissue oxygenation due to impaired microcirculation. In sepsis and septic shock, the microcirculatory alterations also include primary damage to the microcirculation caused by the inflammatory processes and changes to the coagulation system, resulting in a reduced functional capillary density, more non-perfused and intermittently perfused capillaries and an increase in perfusion heterogeneity [3] (Fig. 1D). Other forms of circulatory shock can lead to similar alterations due to secondary damage to the endothelial cells and the tissue [18]. In critically ill patients, altered microcirculation without improvement in disease progression has been shown to be a strong predictor for poor outcome with higher mortality [46]. Measurement of microcirculatory function in critically ill patients with severe hypoxemia and higher SOFA scores due to COVID-19 ARDS showed increased microcirculatory diffusion and convection capacity this in contrast to other viral disease [45, 47, 48]. Representative dark-field microscopy of this population is shown alongside healthy volunteers adapted to high altitude in Fig. 2C. In these patients with isolated lung failure, it was thus possible to study the effects of hypoxemia on an otherwise functionally intact systemic microcirculation and it was shown that adaptation mechanisms to tissue hypoxia are similar to the adaption of healthy volunteers at high altitude. These findings confirm a physiological link between high altitude physiology and critical illness, where in both conditions tissue hypoxia is present. Furthermore, experimental data indicate protective effects associated with adaptation to hypoxia in states of disease, such as a reduced myocardial infarction size in mice when subjected to continuous normobaric hypoxia [49, 50]. These effects show that the intrinsic mechanisms of microcirculation can help the tissue to cope with hypoxemia, provided a sufficient global blood flow and availability of Hb as oxygen carrier.
Hyperoxia may promote microcirculatory dysfunction and cell death through reactive oxygen species (ROS) productionIn contrast to hypoxemia, hyperoxia, defined as excess of oxygen in the tissue and hyperoxemia, being a high blood oxygen content, are often caused by medical staff administering an overabundance of oxygen to the patient. Compared to the macro- and microhemodynamic effects of CO and Hb availability, the effect of differences in oxygen saturation achieved by oxygen supplementation is more difficult to quantify. Hyperoxia induced in the clinical setting by lack of awareness [51] can harm patients through production of ROS and induction of inflammation. At the time of discovery, Joseph Priestley was already discussing possible negative effects of oxygen. Shortly thereafter, Antoine Lavoisier discovered the presence of lung damage in guinea pigs after inhalation of pure oxygen [52]. In 1958, a first report was published on lung damage in humans detected after and possibly related to long-term oxygen therapy [53]. Later research located the main source of ROS within the respiratory chain of the mitochondria in the pulmonary vascular endothelial cells, where the precursor superoxide anion originates at complex III at the inner membrane of mitochondria. The superoxide anion in turn changes into hydrogen peroxide and further turns into water or hydroxyl radicals, which are the main ROS [54]. They are responsible for the adverse effects in tissues across the body. The primary effects of hyperoxia in the lung occur in the form of damage to pulmonary capillary endothelial cells, followed by destruction of pulmonary epithelial cells. Hyperoxia and associated high levels of ROS destroy cellular macro-molecules leading to cell death or initiating apoptosis (Fig. 1D). The effect on remote tissues depends on the inflammatory response with the secretion of chemo-attractants and pro-inflammatory cytokines attracting leukocytes. The leukocytes are thus indirect effectors and at the same time another source of ROS with consecutive inflammation and further destruction of lung and other tissue [55]. High levels of superoxide anions can lead to specific organ damage and ultimately, promote multi-organ failure [56]. The hyperoxic microcirculation primarily shows a decrease in capillary density, that may be accompanied by an increased heterogeneity of capillary perfusion as normally seen in septic patients [13, 57, 58]. Additionally, the mitochondrial oxygen tension (mitPO2) decreased over a level of 26.6 kPa PaO2 [59]. In the systemic vascular bed, hyperoxemia can increase vascular resistance and mean arterial pressure and may decrease CO [60, 61]. Despite this in ovine models of acute peritonitis hyperoxia lead to better macro- and microcirculatory parameters [62]. Whereas a systematic review of hyperoxia in sepsis and septic shock in humans showed in 6 out of 10 included studies an increased mortality [63]. A recent study with mechanically ventilated mice could show time- and dose-dependent immune response of hyperoxia with raised cytokines, neutrophils and chemokines [64]. Knowledge of the relationship between the fraction of inspired oxygen (FiO2) and the formation of ROS particularly above a threshold FiO2 of 0.6 [65], and the mechanisms leading to the adverse effects have increased awareness with oxygen supplementation.
Lower versus higher oxygenation targets in critically ill patientsThe recent advances in our understanding of the effects of both tissue hypoxia and hyperoxia, have underlined the importance of the level of oxygen supplementation not only in terms of a risk–benefit ratio in critically ill patients, but also because of potential protective effects of adaptation mechanisms to hypoxia. Based on the investigation of these pathophysiological mechanisms related to tissue oxygen availability, several clinical studies have been conducted in critically ill patients (Table 1). A trial published in 2014 compared different oxygen saturation (SpO2) targets (SpO2 90–92% versus higher SpO2) and showed only a decrease in lactate levels but no other difference [66]. Another study comparing liberal targets SpO2 above 96% with a conservative group target (SpO2 88–92%) pointed toward a slightly lower 90-mortality in the conservative group [67]. The Oxygen-ICU randomized clinical trial, published in 2016, showed lower ICU-mortality with less episodes of shock, liver failure and bacteremia in the conservative group with an SpO2 target of 94–97% (PaO2 9.3–13.3 kPa) compared to the conventional group with SpO2 of 97–100% (PaO2 up to 20 kPa) [68]. The HYPERS2S-Trial was stopped prematurely when no benefit of hyperoxia with a FiO2 of 1.0 for 24 h compared to a conservative group with SpO2 88–95% could be found [69]. The IOTA review and meta-analysis revealed a dose-dependent increased risk of short- and long-term mortality of patients treated with liberal oxygen [70]. The ICU-ROX investigators found no significant difference in mortality comparing a conservative group with SpO2 < 97% and an usual-oxygen group with no upper limits [71]. On the contrary the LOCO2 Trial was stopped early because of suspicion of an increased risk for serious adverse events and higher 90-day mortality in the conservative group [72]. The biggest prospective study of the HOT-ICU investigators comparing a lower-oxygenation group with PaO2 target of 8 kPa and a higher-oxygenation group with PaO2 of 12 kPa with a total of 2928 patients showed no difference in the 28-day mortality or serious adverse effects [73]. A post hoc subgroup analysis of the cohort did not show any difference in the 90-day mortality between the two groups [74]. Nevertheless, the lower oxygenation group had a significantly higher percentage of days alive without life support. Further a study from the Netherlands with 574 patients (low-normal group PaO2 8–12 kPa, high-normal 14–18 kPa) also found no significant difference in organ dysfunction at 14 days, nor significant differences in 90-day mortality, duration of mechanical ventilation and ICU length of stay [75]. The US PILOT trial, involving 2541 patients, did now show any difference in the number of ventilatory-free days by day 28 between a lower (SpO2 90%), intermediate (SpO2 94%) and a higher (SpO2 98%) oxygenation target group [76]. However, despite the set oxygenation targets, each group in the study
留言 (0)