
記住我
BC is a complex disease at the cellular and molecular levels [11]. Clinical data demonstrate that METTL3 is upregulated in BC tissues, especially in phase T3-T4, or in patients with BC that are diagnosed with tumor metastasis in the lymphatic system [12]. Furthermore, findings also demonstrate the overexpression of METTL3 in BCSCs, and high expression levels of METTL3 are usually associated with a worse prognosis in patients with BC [12]. Therefore, METTL3 might be a risk factor for BC.
METTL3 functions by targeting oncogenesMETTL3-mediated m6A modification of RNAs includes coding (mRNA) and non-coding (i.e., microRNA [miRNA], long non-coding RNA [lncRNA], circular RNA [circRNA]) RNAs.
METTL3 and mRNA modificationBC stem cells (BCSCs) possess self-renewal and differentiation abilities to produce more BC cells, which function primarily during BC initiation, as well as play a fundamental role in BC progression, drug resistance, and recurrence [13]. Xie et al. [12] demonstrated that METTL3 modifies SOX2 mRNA via m6A modification and activates BCSC stemness and malignant progression. Other studies demonstrate that by targeting EZH2 mRNA, METTL3 promotes epithelial-mesenchymal transition (EMT) and metastasis of BCSCs, thereby aggravating the malignant phenotype of BC [14, 15]. In addition, METTL3 also increases the methylation levels, accelerates proliferation rates, promotes tumor growth, and reduces apoptosis of BC cells via m6A modification on Bcl-2 mRNA [14, 15]. Xu et al. [16] reported that METTL3, together with YTHDF2, reduces large tumor suppressor kinase 1(LTSK1) mRNA stability, thereby promoting the proliferation and glycolytic metabolism in BC through the YAP/TAZ axis and Hippo pathways. Furthermore, alternative splicing (AS) enables differential inclusion of exons from a given transcript, thereby contributing to the transcriptome and proteome diversity [17]. Aberrant AS patterns play key roles in BC development. METTL3 functions in modulating BC-associated AS processes [18]. Specifically, both m6A deposition in splice site boundaries and in splicing and transcription factor transcripts, such as MYC, direct AS switches of specific BC-associated transcripts, which are associated with a poor overall survival rate in patients with BC, suggesting the use of these AS events as a novel potential prognostic biomarker [18].
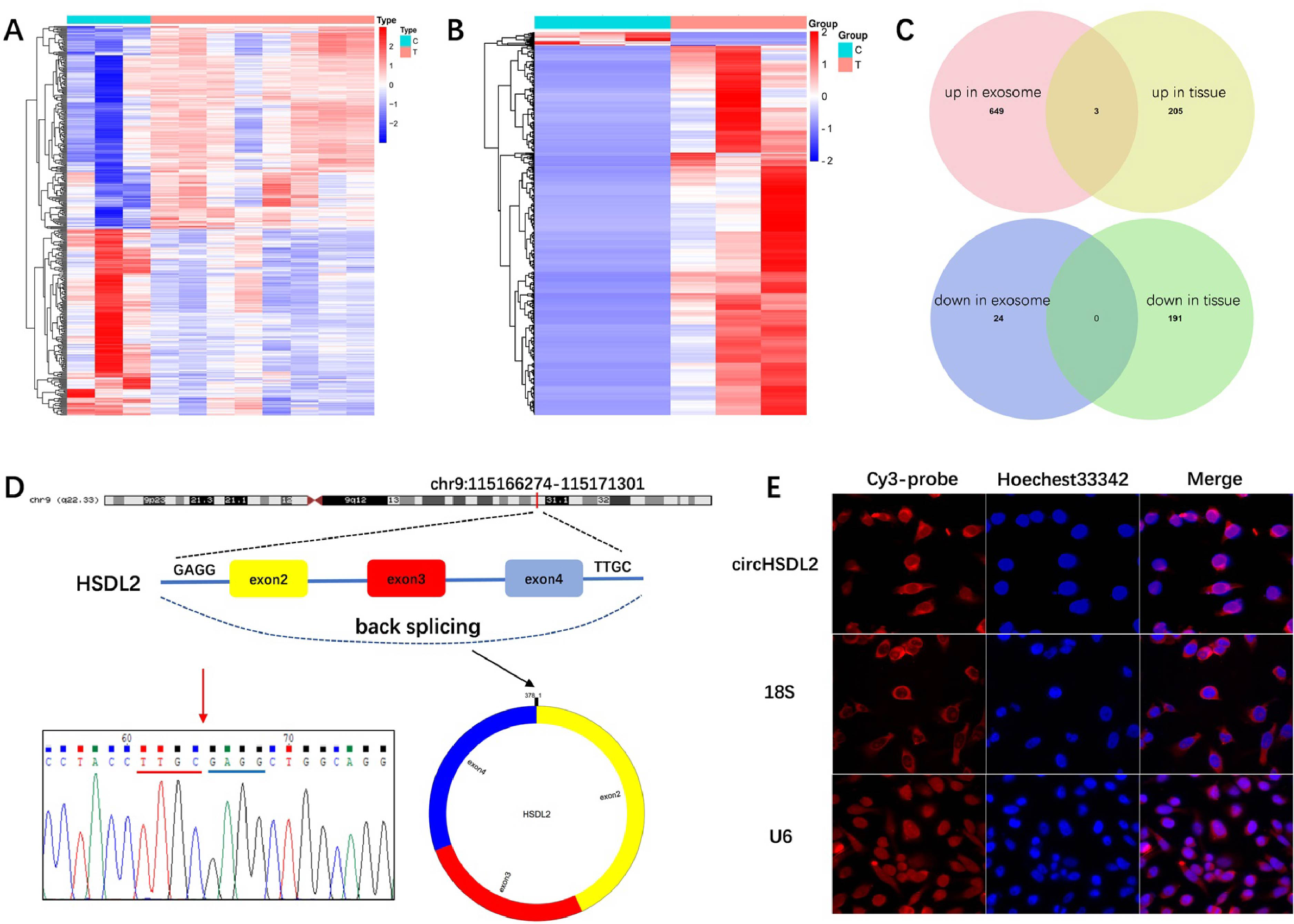
Triple-negative BC (TNBC) is the most dominant and malignant pathological type of BC [19]. m6A regulators exhibit remarkable dysregulation in TNBC tissues, whereas METTL3 shows significant downregulation [20, 21]. Moreover, the level of METTL3 is found to be tightly linked to the progression and poor survival in patients with TNBC, whereas its gene mutation and amplification are not associated with these outcomes [21]. FAM83D is markedly elevated in TNBC tissues and cells, and high-level of FAM83D is related to poor prognosis in patients with TNBC [22, 23]. Yu et al. [22] reported that METTL3 mediates m6A modification on FAM838D and upregulates FAM83D expression to activate the Wnt/β-catenin pathway, which in turn promotes TNBC malignant behaviors [22]. These findings suggest that METTL3 aggravates TNBC progression by targeting the FAM83D/Wnt/β-catenin pathway (Fig. 1).
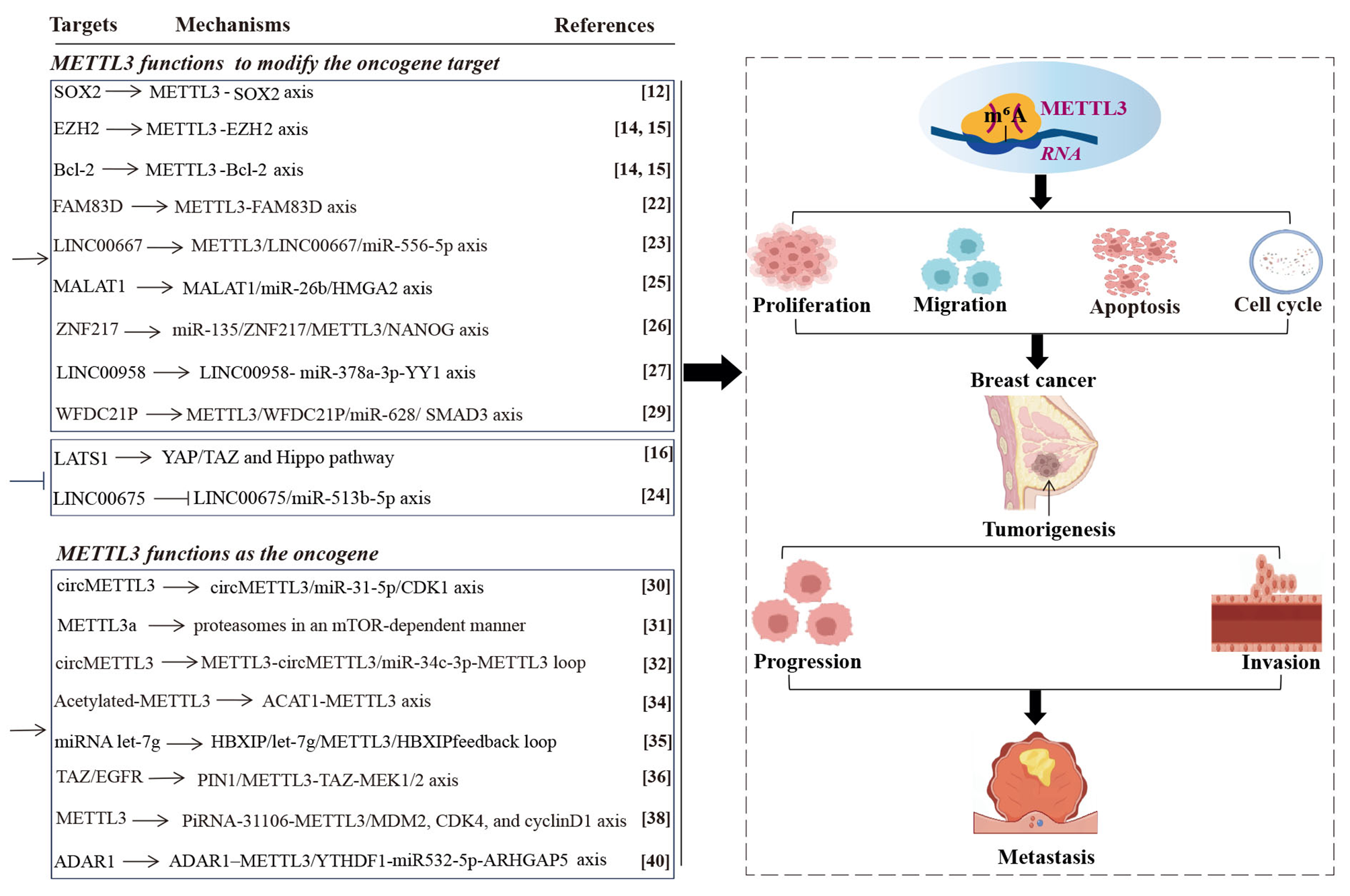
Fig. 1
METTL3-mediated RNA m6A modification induces breast cancer. High levels of METTL3 are associated with the tumorigenesis, progression, invasion, and metastasis of breast cancer, which functions by mediating m6A modification on different RNAs, inducing the abnormal process of proliferation, migration, apoptosis, and cell cycle. Methyltransferase-like 3 (METTL3). N6-methyladenosine (m6A). Acetyl-CoA acetyltransferase 1 (ACAT1). Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1). High mobility group AT-hook 2 (HMGA2). High zinc finger protein 217 (ZNF217). Large tumor suppressor kinase 1 (LATS1). Transcriptional coactivator with PDZ-binding motif (TAZ). Host gene of circular RNA METTL3 (circMETTL3). Cyclin-dependent kinase 1 (CDK1). Cleaved form of METTL3 (METTL3a). Hepatitis B X-interacting protein (HBXIP). Epidermal growth factor receptor (EGFR). Peptidyl-prolyl cis-trans isomerase NIMA-interacting 1 (PIN1). P-element-induced wimpy testis (PIWI)-interacting RNAs (piRNAs). YTH N6-methyladenosine RNA binding protein F1 (YTHDF1)
Collectively, these findings suggest that METTL3 functions as an oncogenic driver to modify mRNAs and triggers BC onset and progression.
METTL3 and non-coding RNAsMETTL3 also exerts its oncogenic function by modulating the expression of noncoding RNAs. Ren et al. [23] found that LINC00667 is an m6A-modified lncRNA, which is up-regulated upon KIAA1429 overexpression. The high expression of LINC00667 is correlated with the poor prognosis of patients with BC. LINC00667 promotes the proliferation and migration of BC cells. KIAA1429 targets the m6A modification site on LINC00667 and enhances its mRNA stability [24]. Moreover, LINC00667 positively regulates KIAA1429 via sponging miR-556-5p, forming a KIAA1429/m6A/LINC00667/miR-556-5p feedback loop [24]. Collectively, the central findings of this study suggest that KIAA1429-induced LINC00667 exerts functions as an oncogene in BC progression through an m6A-dependent feedback loop. miR-26b exhibits consistently low levels, while METTL3, metastasis-associated lung adenocarcinoma transcript 1 (MALAT1), and high mobility group AT-hook 2 (HMGA2) demonstrate elevated levels in BC [25]. METTL3 increases MALAT1 expression by modulating its m6A modification, which subsequently promotes HMGA2 expression through miR-26b sponging, ultimately leading to EMT, migration, and invasion of BC cells [25]. Notably, METTL3 drives BC tumorigenesis via the MALAT1/miR-26b/HMGA2 axis.
High zinc finger protein 217 (ZNF217) expression and low miR-135 expression levels are found in BC tissues and cells [26]. Silencing ZNF217 or elevating miR-135 inhibits BC cell malignant behaviors [26]. ZNF217 upregulates METTL3 and that in turn targets NANOG to promote BC progression [26]. Therefore, the miR-135/ZNF217/METTL3/NANOG axis promotes BC progression, emphasizing METTL3 or NANOG as a potential therapeutic target for BC treatment. Rong et al. [27] showed that METTL3 facilitates LINC00958 expression by enhancing its stability in BC tissues and cells, thus revealing a novel METTL3-involving mechanism for regulating BC tumorigenesis. LINC00958 interacts with miR-378a-3p in a competitive endogenous RNA (ceRNA) manner to promote YY1 expression [27]. Therefore, BC tumor progression is further promoted and aggravated via the METTL3-LINC00958/miR-378a-3p-YY1 axis. Fan et al. [28] discovered that METTL3 downregulates LINC00675 expression, which is indicative of a higher tumor grade, worse lymphovascular invasion, and shorter survival in patients with BC. Mechanistically, LINC00675 acts as a ceRNA by interacting with miR-513b-5p and repressing its expression [28].
Furthermore, the level of LncRNA-WFDC21P is abnormally high in TNBC, which promotes TNBC cell proliferation and metastasis, as well as is strongly linked to a poor survival rate of patients. Wei et al. [29] reported that METTL3-mediated m6A modification on WFDC21P upregulates WFDC21P expression in TNBC cells, which in turn interacts with miR-628-5p and further inhibits Smad3-related gene expression to promote cell proliferation and metastasis [29]. This study demonstrates the vital roles of METTL3-mediated m6A modification on WFDC21P and its regulation in the proliferation and metastasis of TNBC cells via the METTL3/WFDC21P/miR-628/SMAD3 axis (Fig. 1).
METTL3 functions as the target oncogene METTL3 functions in BC by modifying itselfIn addition to regulating target gene expression, METTL3 itself is regulated by other factors in BC. Li et al. [30] revealed that METTL3, the host gene of circular RNA METTL3 (circMETTL3), regulates circMETTL3 expression in an m6A-dependent manner. However, circMETTL3 does not affect METTL3 expression. Mechanistically, circMETTL3 interacts with miR-31-5p in a ceRNA manner, thereby upregulating the expression of its target, cyclin-dependent kinase 1 (CDK1) [30]. Therefore, circMETTL3 facilitates BC cell proliferation, migration, and invasion via the circMETTL3/miR-31-5p/CDK1 axis. In addition to investigating the novel relationship between the circRNA and its corresponding host gene, another study indicated that the cleaved form of METTL3 (termed METTL3a), which is mediated by proteasomes in an mTOR-dependent manner, is essential for regulating the assembly of the METTL3-METTL14-WTAP complex [31]. Subsequently, the METTL3-METTL14-WTAP complex functions to promote BC progression by depositing m6A modification on RNAs, which primarily depends on its methyltransferase activity that is supported by the catalytic role of METTL3, the structural role of METTL14, as well as the adaptor role of WTAP.
In contrast, a previous study revealed a striking reduction of METTL3 levels in TNBC samples, including tissues and cell lines [32]. Functionally, METTL3 upregulates its target, circMETTL3, which consequently interacts with miR-34c-3p in a ceRNA manner [32]. Furthermore, miR-34c-3p represses METTL3 expression [32]. The negative feedback-loop regulatory mechanism of METTL3-circMETTL3/ miR-34c-3p-METTL3 promotes TNBC tumor cell proliferation, invasion, metastasis, and growth [32]. Meanwhile, a low level of METTL3 in TNBC is strongly linked to short, distant metastasis-free survival [33]. METTL3 augments migration, invasion, and adhesion of TNBC cell lines by upregulating the collagen type III alpha 1 chain, ultimately promoting the metastasis of TNBC cells [33]. In addition, Zhang et al. [34] reported an acetylation-related mechanism for TNBC progression, demonstrating that nuclear receptor subfamily 2 group F member 6 transcriptionally activates acetyl-CoA acetyltransferase 1 (ACAT1), which promotes TNBC cell migration and invasion via ACAT1-mediated METTL3 acetylation. Notably, although the level of METTL3 is downregulated, the modified-METTL3 (circMETTL3, METTL3 acetylation) is upregulated, which together affect the malignant processes in BC (Fig. 1).
Collectively, these findings suggest that METTL3 contributes to BC by regulating itself, suggesting its oncogenic role. Moreover, although findings suggest a low level of METTL3 in TNBC, it functions through its modified types with the oncogenic role. Therefore, METTL3 is a risk gene for BC.
METTL3 functions through a regulatory networkConsistently, Cai et al. [35] reported that hepatitis B X-interacting protein (HBXIP) increases METTL3 expression in BC by suppressing miRNA let-7 g. Remarkably, METTL3-mediated m6A modification in turn enhances HBXIP expression [35]. Overexpression of HBXIP not only facilitates BC cell proliferation but also impairs apoptosis [35]. In summary, a positive feedback-loop regulatory mechanism, HBXIP-let-7 g/METTL3-HBXIP, is established by HBXIP to expedite BC cell proliferation. Peptidyl-prolyl cis-trans isomerase NIMA-interacting 1 (PIN1) interacts with METTL3 and thereby blocks its ubiquitin-dependent proteasomal and lysosomal degradation [36]. In clinical settings, METTL3 expression is significantly increased with tumor progression and is positively correlated with PIN1 expression in BC tissues [36]. PIN1 stabilizes METTL3, which in turn increases the m6A modification and translation of the transcriptional coactivator with PDZ-binding motif (TAZ) and the epidermal growth factor receptor (EGFR) mRNA [36]. Inhibition of MEK1/2 kinases and PIN1 destabilizes METTL3, which blocks BC cell proliferation and induces cell cycle arrest at the G0/G1 phases [36]. Moreover, silencing METTL3 reduces PIN1 overexpression-induced colony formation in MCF7 cells and enhances tumor growth in 4T1 cells in mouse models [36]. Therefore, PIN1 upregulates METTL3, and the PIN1/METTL3 axis may be an alternative therapeutic target for BC. P-element-induced wimpy testis (PIWI)-interacting RNAs (piRNAs) are novel non-coding RNAs whose abnormal expression is closely associated with multiple cancers [37]. Huang et al. [38] reported that the level of PiRNA-31,106 is high in BC tissues and cell lines (MDA-MB-231 and MCF-7), which promotes the m6A methylation levels and facilitates METTL3 expression that further promotes BC progression by upregulating the expressions of MDM2, CDK4, and cyclinD1 [38]. Therefore, PiRNA-31,106 upregulates METTL3 to promote BC progression, suggesting its potential as a therapeutic target in BC (Fig. 1).
To sum up, these findings highlight that METTL3 regulates BC progression through various mechanisms, including interacting with other molecules and influencing multiple pathways. Therefore, while METTL3 does have a direct impact on oncogenic processes, its role is also mediated through complex regulatory networks.
METTL3 functions by interacting with A-to-I RNA editingFurthermore, A-to-I RNA editing is another prevalent type of RNA modification that controls gene expression in mammals and regulates tumorigenesis and tumor progression [39]. Specifically, ADAR1 is an A-to-I RNA-editing enzyme, both ADAR1 and METTL3 are upregulated in BC samples [40]. Li et al. [40] showed that ADAR1 interacts with METTL3 in a YTHDF1-dependent manner and further increases METTL3 expression. Mechanically, ADAR1 changes the binding site of METTL3 mRNA on miR532-5p and is recognized by YTHDF1. Subsequently, METTL3 expression is increased, which further targets ARHGAP5 to promote the proliferation, migration, and invasion of BC cells [40]. Loss of ADAR1 or METTL3 significantly inhibits BC growth in vivo [40]. Notably, the ADAR1-METTL3 axis is a novel pathway that connects the RNA regulation mechanism of A-to-I editing and METTL3-mediated m6A modification during BC progression. Therefore, the combined targeting of ADAR1 or METTL3 might be more effective for BC treatment (Fig. 1).
Collectively, these findings suggest that METTL3 is primarily associated with BC initiation and progression, which functions by regulating the oncogene target or functions as the target oncogene. Furthermore, these findings suggest that METTL3 has great clinical potential as a diagnosis biomarker.
METTL3 induces drug resistance and poor prognosis of BCChemotherapy is an important strategy for BC treatment [41]. Adriamycin (ADR), Tamoxifen, and Paclitaxel are currently considered the primary therapeutic drugs for patients with BC [42]. However, clinical data suggest that the majority of patients with BC experience resistance to chemotherapy [41]. Specifically, resistance to ADR and Tamoxifen is considered a troublesome challenge for curing patients with BC [43]. Currently, studies have also suggested the implication of METTL3-mediated RNA m6A modification in the development of BC chemoresistance. The global levels of mRNA m6A methylation and METTL3 are upregulated in ADR-resistant BC cell lines while silencing METTL3 partially overcomes this resistance [44]. METTL3 cooperates with IGF2BP3 to modulate the m6A modification on HYOU1 mRNA and increases HYOU1 stability, which subsequently increases ADR resistance in BC cells [44]. Therefore, the METTL3/IGF2BP3-HYOU1 axis modulates ADR sensitivity in BC cells, targeting this axis might be a strategy to improve ADR efficacy in BC treatment.
Zhou et al. [43] demonstrated that METTL3-mediated m6A modification on long noncoding RNA (lncRNA) KCNQ1OT1 promotes ADR resistance in BC by regulating the miR-103a-3p/multidrug resistance protein 1 (MDR1) axis. LncRNA KCNQ1OT1 is highly expressed in ADR-resistant BC cells, and its expression is modulated by MELLT3 via m6A modification. A further mechanistic study found that miR-103a-3p acts as a sponge for lncRNA KCNQ1OT1 and MDR1 [43]. Altogether, these findings indicate that the METTL3-lncRNAKCNQ1OT1/ miR-103a-3p-MDR1 axis is responsible for ADR resistance in BC, which suggests a novel approach to prevent ADR resistance in patients with BC. Consistently, high levels of METTL3 enhance the ADR resistance for BC cells by upregulating miR-221-3p through positive regulation of homeodomain-interacting protein kinase 2 (HIPK2) and its direct target Che-1 [45]. More detailed, METTL3 functions to promote miR-221-3p expression by decreasing m6A mRNA methylation on pri-miR-221-3p [45]. Subsequently, miR-221-3p further enhances the expression of MDR1 and BCRP, which promotes a higher production of ADR-resistant cells while inhibiting apoptosis in such cells [45]. Together, after the cooperation of these signaling molecules, the METTL3-HIPK2/Che-1-miR-221-3p-MDR1/BCRP axis triggers ADR resistance in BC cells. Li et al. [46] found that METTL3 is highly expressed in ADR-resistant BC cells and upregulates MALAT1 through m6A modification, which, in turn, recruits E2F transcription factor 1(E2F1) to activate anterior gradient 2 (AGR2) transcription and increases ADR resistance in BC cells. Collectively, METTL3 promotes ADR resistance in BC by targeting the MALAT1/E2F1/AGR2 axis. Moreover, Li et al. [47] reported that METTL3 promotes the efficiency of homologous recombination repair and inhibits ADR-induced DNA damage. METTL3 functions by enhancing epidermal growth factor (EGF) synthesis in a YTHDC1-dependent manner, and EGF subsequently inhibits RAD51 expression [47]. These findings reveal the mechanism of METTL3 in regulating ADR resistance in BC by targeting the EGF/RAD51 axis. Furthermore, Liu et al. [48] revealed that the expression of METTL3 and adenylate kinase 4 (AK4) is remarkably elevated in Tamoxifen-resistant BC cells compared with that in their parental counterparts. Further investigation found that METTL3 methylates AK4 mRNA on the 5′-untranslated region, leading to Tamoxifen resistance in patients with BC [46].
Consistently, Jing et al. [49] reported that METTL3 regulates Docetaxel resistance of TNBC cells by regulating a feedback loop. They found that METTL3 uses LINC00662 as its target, which directly interacts with miR-186-5p, and METTL3 expression is in turn regulated by miR-186-5p [49]. LINC00662 and miR-186-5p regulate the viability of Docetaxel-resistant BC cells, and miR-186-5p subsequently regulates METTL3 expression [49]. Overall, these findings suggest that suppression of METTL3 promotes Docetaxel resistance in BC through the METTL3-LINC00662/miR‑186‑5p-METTL3 signaling (Fig. 2).
Fig. 2
METTL3 contributes to the drug resistance of breast cancer. High levels of METTL3 are associated with drug resistance and poor prognosis of breast cancer by mediating m6A modification on different RNAs. Methyltransferase-like 3 (METTL3). N6-methyladenosine (m6A). Multidrug resistance protein 1 (MDR1). Homeodomain-interacting protein kinase 2 (HIPK2). Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1). E2F transcription factor 1(E2F1). Anterior gradient 2 (AGR2). Epidermal growth factor (EGF). YTH domain containing 1 (YTHDC1). Adenylate kinase 4 (AK4)
However, METTL3 knockdown decreases the drug sensitivity of hormone receptor-positive/human epidermal growth factor receptor 2-negative (HR + HER2−) cells by targeting CDKN1A, activating the AKT pathway, and promoting EMT [50]. Moreover, METTL3 knockdown also leads to the inhibition of BAX and caspase-3/-9/-8 [50]. Therefore, METTL3 might play a tumor-suppressor role and it could be a potential biomarker for predicting the prognosis of patients with HR + HER2-BC.
Collectively, these data suggest that METTL3 contributes to various drug resistance of BC, which usually indicates a poor prognosis and recurrence of BC. Therefore, we suppose that METTL3 is a promising prognostic biomarker for BC.
Targeting METTL3 for BC therapyConsidering that a high level of METTL3 promotes BC occurrence, progression, and drug resistance. Therefore, inhibition of METTL3 is considered a potential approach for promoting chemotherapeutic effectiveness against BC. STM2457, the small-molecule inhibitor of METTL3, significantly reduces the proliferation, viability, and migration of TNBC cells, including both BRCA1/2 wild-type and BRCA1-mutated cell lines [51]. Therefore, STM2457 displays anti-tumor activity in TNBC and enhances chemotherapy response while sensitizing BRCA1/2 wildtype patients to Olaparib.
Epigenetic drugs are a novel therapeutic method for BC [52]. Cromolyn is a mast cell stabilizer emerging as an anticancer drug [
留言 (0)