
記住我
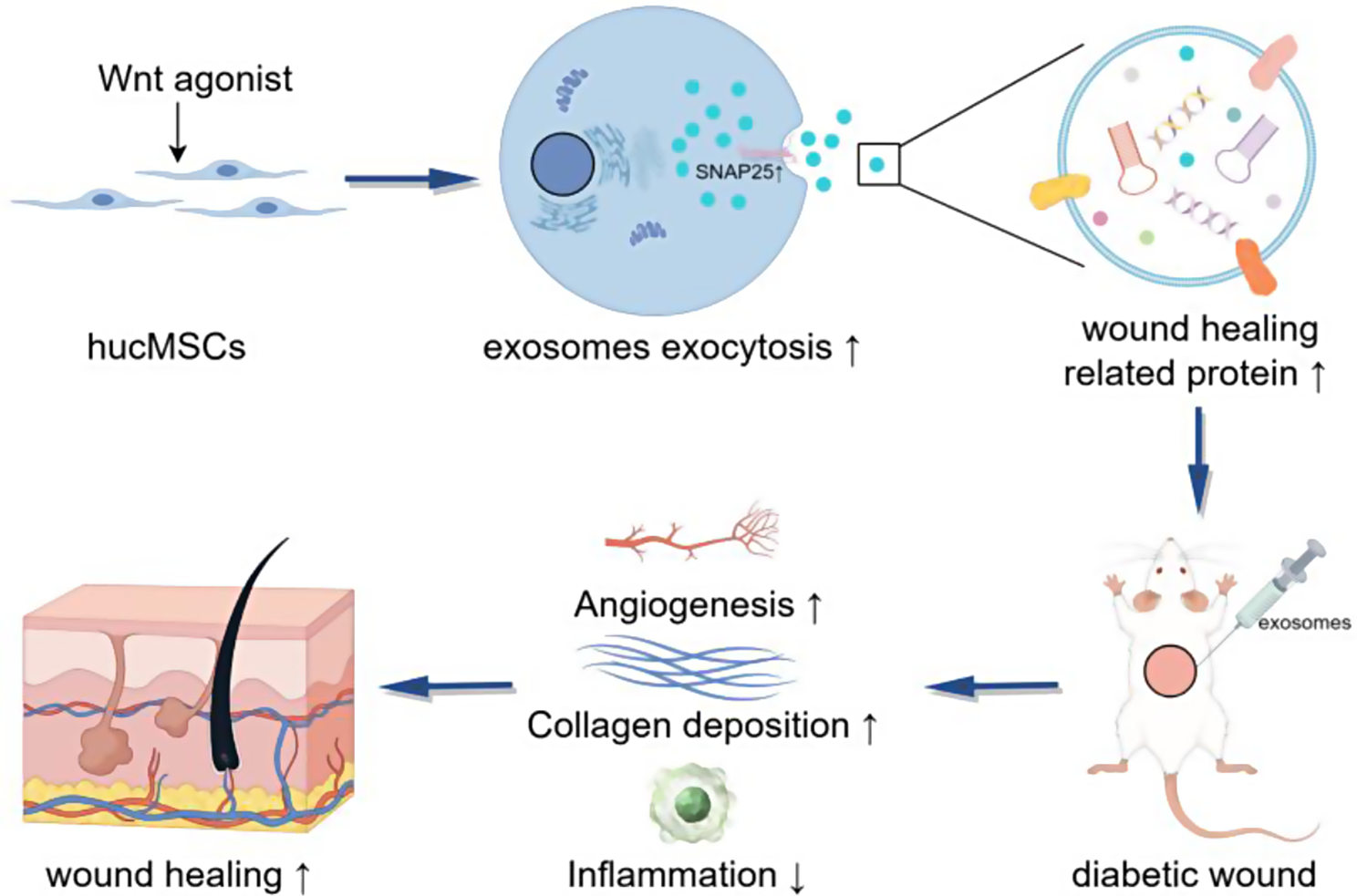


Before examining the impact of the Wnt signalling pathway on the production and cargo loading of exosomes, we had to verify that the properties of hucMSCs and exosomes after Wnt signalling priming were unaltered. In this study, 2 Wnt agonists (CHIR99021 and CP21R7) and 1 inhibitor (XAV939) were tested. After treatment, the ability of the pretreated hucMSCs to differentiate into adipocytes or osteocytes did not significantly change compared to that of the untreated hucMSCs, suggesting that altered Wnt signalling had no obvious effect on the differentiation potential of MSCs (Fig. 1A). In addition, flow cytometry analysis revealed that the hucMSCs subjected to different treatments exhibited comparable negative CD34 and HLA-DR expression and positive CD105 and CD73 expression, suggesting that these procedures did not significantly change the surface marker profile of the MSCs (Fig. 1B). Furthermore, to evaluate cell senescence, we cultured hucMSCs with or without CHIR99021 pretreatment for 7 continuous passages. By the 7th passage, the β-galactosidase-positive area was significantly reduced by priming with CHIR99021 compared to that in the untreated hucMSCs, suggesting that CHIR99021 delayed the senescence of hucMSCs (Figure S1A).
Exosomes were then isolated from hucMSCs that were subjected to different pretreatment procedures and characterized via Western blotting analysis. The results revealed the presence of TSG101, CD63, and CD9 in the exosomes from all four groups, while calnexin, which is a negative marker of exosomes, was absent (Fig. 1C). TEM analysis confirmed the characteristic double-membrane structure that is commonly observed in exosomes (Fig. 1D). Finally, the size distribution of the exosome particles was determined using NTA analysis, and the results indicated an average particle size of approximately 130 nm in the exosomes of all four groups (Fig. 1E). These findings collectively indicated that the administration of Wnt signalling pathway agonists or inhibitors does not exert any discernible effect on the characteristics of MSCs or the inherent properties of the secreted exosomes.
Fig. 1
The characteristics of hucMSCs and exosomes remained unchanged after Wnt signalling priming. (A) Alizarin red and oil red O staining after treatment of hucMSCs with Wnt agonists or inhibitors (B) Flow cytometry identification of hucMSCs maker after treatment of hucMSCs with Wnt agonists or inhibitors. (C) Western blot analysis of exosome marker from different pretreated hucMSCs. (D) Exosome transmission electron microscopy. (E) Exosome NTA assay
Wnt agonist treatment increased hucMSC exosome productionSubsequently, we investigated the potential impact of the Wnt signalling pathway on exosome production in hucMSCs. Previous studies demonstrated a positive correlation between exosome content and AChE levels in culture supernatants [24]. Therefore, we initially quantified the AChE concentration in the MSC supernatant as an indicator of exosome content. To ensure accurate comparisons, we normalized the results according to the total protein content. A 1.5-fold increase in AChE levels was observed in the groups treated with Wnt signalling pathway agonists, whereas inhibition of this pathway led to a decrease in exosome secretion (Fig. 2A). To further verify this trend, we employed NTA to quantify the exosome size and number. The exosome number was normalized according to the number of MSCs. The NTA results proved that treatment with Wnt agonists resulted in a similar 1.5-fold increase in exosome secretion (Fig. 2C). Notably, this increase in secretion did not significantly influence the particle size of the exosomes (Fig. 2B). We also found that the effect of the agonist CHIR99021 was slightly more pronounced than that of CP21R7 after the results were normalized to the cell number, but the difference was not statistically significant (Fig. 2B-C). Additionally, we utilized TEM to visualize the intracellular content of the MVBs and intraluminal vesicles (ILVs). Our observations revealed an increase in the intracellular MVBs and ILVs following treatment with a Wnt signalling pathway agonist (Fig. 2D-F). Collectively, the above results suggested that Wnt signalling pathway agonists can increase the content of MVBs and ILVs within hucMSCs and increase exosome secretion by approximately 1.5-fold. Nevertheless, considering the consistency of the results, we chose CHIR99021 as the Wnt agonist for use in the subsequent proteomic, transcriptomic and functional validation experiments.
Fig. 2
Wnt agonist treatment increased hucMSC exosome production. (A) ELISA detection of AChE in hucMSC supernatants from different treatment groups (B) Exosome particle size distribution. (C) Quantification of different treatment groups of exosomes by NTA. (D-F) Representative TEM images of hucMSCs and the quantitation analysis on the MVBs and ILVs after hucMSCs were treated with Wnt agonists or inhibitors. Data are represented as mean ± SEM. *P < 0.05, **P < 0.01
CHIR99021 increased exosome secretion by promoting SNAP25-dependent exocytosisOur aforementioned results suggested that Wnt signalling pathway agonists can increase exosome production by 1.5-fold. Previous studies revealed that exosome formation is affected mainly through the following three key processes: MVB and ILV production, lysosome-mediated degradation, and exocytosis.
To investigate the underlying mechanisms driving this enhanced secretion, we conducted transcriptomic analysis of hucMSCs treated with or without CHIR99021. Differentially up- and downregulated genes were revealed by volcano plot analysis (Fig. 3A). Subsequent Gene Ontology (GO) analysis indicated that the differentially expressed genes were enriched in the term “extracellular exosomes” (Fig. 3B). Furthermore, KEGG analysis revealed potential enrichment in the “synaptic vesicle cycle” pathway, which is a crucial pathway involved in cytosolic exocytosis (Fig. 3C). By conducting a clustering heatmap analysis of the 35 genes associated with the synaptic vesicle cycle, we found that 24 genes were upregulated in the CHIR99021 group. Notably, one of the upregulated genes was SNAP25, an essential constituent of the soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) complex that plays a critical role in exocytosis processes (Fig. 3D). To validate this result, we conducted Western blotting analysis and observed a similar upregulation of SNAP25 protein expression induced by CHIR99021. Additionally, CHIR99021 upregulated the expression of exocytosis regulatory proteins, namely, Rab27a and Rab7a. Rab27a promotes the accumulation and transport of MVBs to the plasma membrane, while Rab7a mediates MBV maturation and transport and endosome-lysosome fusion. Conversely, treatment with the Wnt signalling inhibitor XAV939 had the opposite effect (Fig. 3E). Notably, no changes were observed in the protein levels of other SNARE complex components, including STXBP1 VAMP2 and VAMP3, in the expression of the ESCRT components ALIX and TSG101 or in the lysosome marker LAMP2 (Figure S1B). Taken together, these results demonstrated that Wnt signalling regulates the exocytosis of hucMSCs.
Next, we utilized TEM and confocal microscopy to visualize the intracellular localization and content of MVBs and ILVs. Considering that SNAP25 interacts with its ligand VAMP3 to trigger membrane fusion and exosome release, we first examined the colocalization of SNAP25 and VAMP3. SNAP25 and VAMP3 were significantly colocalized near the plasma membrane in the Wnt agonist-treated group (Fig. 3F), suggesting that the Wnt agonist increased SNAP25-mediated membrane fusion. Additionally, RAB7a, which mediates MBV maturation and transportation, exhibited increased fluorescence enrichment at the cytosolic membrane after agonist treatment (Figure S1C). Regarding lysosome-mediated degradation, the colocalization of the lysosome marker LAMP2 with the MVB marker CD63 was reduced after Wnt agonist treatment, suggesting that the lysosome-mediated degradation of MVBs might be suppressed by the Wnt agonist (Fig. 3G). These findings were consistent with our TEM results, which showed an increase in the number of intracellular MVBs and ILVs after agonist priming (Fig. 2D-F). No significant changes were observed in the levels of exosome synthesis-related proteins, such as ALIX and TSG101 (Figure S1B). The Wnt agonists may have increased the MVB and ILV contents by suppressing lysosome-mediated degradation. However, we cannot exclude the possibility that Wnt agonists might regulate the generation of MVBs and ILVs through mechanisms that are independent of ALIX and TSG101, which needs further verification. We will only focus on the mechanism validated by transcriptomics, namely, the SNAP25-mediated exocytosis pathway.
To demonstrate whether the regulatory effect of Wnt agonists on exocytosis is SNAP dependent, we used siRNA to knock down SNAP25 (Fig. 3H). Given the consistent results observed in Fig. 2 between the AChE level analysis and NTA, we measured AChE levels as an indicator of exosome content. Notably, suppression of SNAP25 expression resulted in a significant reduction in AChE levels, and the pro-exocytosis effects of the Wnt agonists were nearly abolished (Fig. 3I). These findings suggested that Wnt signalling pathway agonists can potentially facilitate exosome release, and the underlying mechanism is partially dependent on SNAP25.
Fig. 3
CHIR99021 increased exosome secretion by promoting SNAP25-dependent exocytosis. (A) Volcano plot showing differential gene expression of hucMSCs in the CHIR99021-treated group versus the control group. (B) GO enrichment analysis of hucMSCs in CHIR99021-treated group versus control group. (C) KEGG enrichment analysis of hucMSCs in CHIR99021-treated group versus control group. (D) Heatmap of gene clustering of hucMSCs transcriptomics in synaptic vesicle cycle between CHIR99021-treated and control groups. (E) Western blot analysis of β-catenin, Rab27a, Rab7, SNAP25 in hucMSCs treated with Wnt signaling pathway agonists and inhibitors. (F) Co-location of SNAP25 with VAMP3 was observed using immunofluorescence after treatment of hucMSCs with Wnt signaling pathway agonists and inhibitors. (G) Co-location of LAMP2 with CD63 was observed using immunofluorescence after treatment of hucMSCs with Wnt signaling pathway agonists and inhibitors. (H) Western blot analysis of SNAP25 knockdown efficiency. (I) After knocking down of SNAP25, hucMSCs were treated with Wnt signaling pathway agonists for 48 h, supernatants were collected, and AChE concentration was analysis by ELISA. Data are represented as mean ± SEM. *P < 0.05, **P < 0.01
CHIR99021 enhanced the loading of wound healing-related proteins into exosomesBased on the above results, Wnt agonists, particularly CHIR99021, significantly increased SNAP25-mediated exosome secretion. However, importantly, an increase in exosome production alone cannot guarantee therapeutic efficacy. Therefore, we aimed to explore the impact of Wnt agonists on the cargo content of exosomes by conducting proteomics analysis of exosomes that were released by hucMSCs with or without CHIR99021 treatment (referred to as CHIR-Exos and CTRL-Exos, respectively). The volcano plot shows the differentially upregulated and downregulated proteins in response to CHIR99021 treatment (Fig. 4A). GO analysis revealed that the proteins exhibiting altered expression were enriched in two biological processes, namely, “positive regulation of cell migration” and “wound healing” (Fig. 4B). A clustered heatmap of the proteins enriched in these processes highlighted the upregulation of TGFβ2, TGFβ3, AGO2, CCAR1 and HMGB1, which are known to promote wound healing and cell migration (Fig. 4C-D). Transcriptomic analysis of hucMSCs revealed similar results, with differentially expressed mRNAs clustering in the TGFβ pathway and PI3K-Akt pathway, both of which play important roles in regulating wound healing (Fig. 4E-F). These results indicated that CHIR99021 treatment could alter the contents of exosomes and promote the enrichment of proteins that are associated with wound healing and cell migration.
Fig. 4
CHIR99021 enhanced the loading of wound healing-related proteins into exosomes. (A) Volcano plot showing differential protein of exosome between CHIR99021-treated and control groups. (B) GO enrichment analysis between CHIR99021 treatment group and control group. (C-D) Heatmap showing differential protein between CHIR99021 treatment group and control group. (E-F) Heatmap of hucMSCs transcriptomics clustering in CHIR99021-treated group vs. control group
CHIR-Exos promoted diabetic cutaneous wound healing more strongly than untreated hucMSC exosomesTo further confirm whether the CHIR-Exos could promote wound healing in vivo, we established a full-thickness cutaneous wound model in diabetic mice (Fig. 5A). Subsequent imaging on days 0, 3, 7, 10, and 14 revealed that CHIR-Exos significantly accelerated the healing of diabetic wounds, surpassing the healing rate induced by control exosomes (referred to as CTRL-Exos). Notably, the CHIR-Exo group achieved near-complete wound closure by the 14th day of observation (Fig. 5B-D).
Histological analysis of skin sections from wounds on day 14 revealed a notable increase in the epidermal layer in the CTRL-Exo treatment groups. Remarkably, this increase was even more pronounced in the CHIR-Exo treatment group, in which the cells were arranged in a more normal pattern (Fig. 5E-F). Since Exos have been reported to engage in and enhance nearly all stages of wound healing [25], immunohistochemical staining and Masson staining were performed to assess collagen deposition, the inflammatory response, and angiogenesis in wounds. The results indicated that the deposition of both collagen I and collagen III was enhanced in the wounds of the CHIR-Exo group, and this change was accompanied by significantly reduced IL1β expression and enhanced angiogenesis at the wound site (Fig. 5G-K). These findings indicated that the administration of CHIR-Exos could enhance collagen deposition, suppress the inflammatory response and facilitate angiogenesis in diabetic wounds, thereby promoting the process of wound healing.
Fig. 5
CHIR-Exos promoted diabetic cutaneous wound healing more strongly than untreated hucMSC exosomes. (A) Flow chart of the animal experiment. (n = 5 in each group) (B) Skin wound healing in 4 groups of mice on days 0, 3, 7, 10 and 14. (C) Pattern diagrams of skin healing in the 4 groups of mice, with coloured areas showing the area of unhealed wounds at different observation times. (D) Statistics on the percentage of wound healing area of mice. (E-F) H&E staining to observe the epidermal thickness of the skin at the wound in each group of mice. (G-K) Masson staining and immunohistochemical analysis of collagen deposition, IL-1β, CD31 expression in the skin of mice in each group. Data are represented as mean ± SEM. *P < 0.05, **P < 0.01
CHIR-Exos promoted fibroblast migration and proliferation and suppressed apoptosis in vitroWe further verified the pro-healing effects of CHIR-Exos in the NIH/3T3 cell line. Different exosome treatments were administered to stimulate cell migration, validating the effects of exosomes from various groups on this process. The uptake of all three types of exosomes by HIN/3T3 cells was confirmed (Figure S1D). Notably, both the CTRL-Exos and CHIR-Exos significantly promoted HIN/3T3 cell migration, with the CHIR-Exos exhibiting a stronger effect than the CTRL-Exos. Conversely, XAV939-pretreated exosomes (referred to as XAV-Exos) exerted the weakest promotion of cell migration (Fig. 6A-B). Annexin V/PI staining demonstrated that the CHIR-Exos inhibited apoptosis (Fig. 6C), while the cell cycle assay revealed that these exosomes also increased the proportion of cells in the S and G2 phases, thereby promoting cell proliferation (Fig. 6D). Consistent with the Annexin V/PI staining results, CHIR-Exo treatment also decreased the protein level of cleaved caspase 3 (Fig. 6F) as well as the activation of the PI3K/AKT signalling pathway and the P38MAPK signalling pathway. KEGG analysis revealed similar results; namely, the differentially expressed proteins in the CHIR-Exos were significantly enriched in the PI3K/AKT signalling pathway and cell adhesion molecules (Fig. 6E). These findings indicated that the CHIR-Exos promoted cell migration, suppressed oxidative stress-induced apoptosis, and prevented cell cycle arrest in vitro.
Fig. 6
CHIR-Exos promoted fibroblast migration and proliferation and suppressed apoptosis in vitro. (A-B). Migration of NIH/3T3 cells at 0, 6, 12, and 24 h in scratch assay. (C) Apoptosis was detected in each group using Annexin5/PI staining, and the histogram showed the percentage of both Annexin5 (+) PI (−) and Annexin5 (+) PI (+) cells. (D) Cell cycle was detected using PI staining. (E) KEGG enrichment analysis of exosome between CHIR99021-treated and control groups. (F) Western blot analysis of AKT, p-AKT, p38, p-p38, caspase3 and cleaved-caspase3 in each group. Data are represented as mean ± SEM. *P < 0.05, **P < 0.01
Discussion.
MSCs are a promising therapy for various diseases ranging from diabetes to wound healing [26, 27]. Exosomes are small EVs with a size of 30–150 nm and exert most of the therapeutic effects of MSCs. Exosomes transport a diverse range of molecules, including proteins, mRNAs, miRNAs, and lipids, and thus play crucial roles in various biological processes, such as intercellular communication, immunomodulation, tumour metastasis and tissue regeneration [28,29,30]. However, the poor scalability of their production has limited the large-scale application of MSC-derived exosomes in the clinic. Therefore, various approaches have been developed to increase their production and reduce their heterogeneity [31]. These approaches range from new cell culture devices to priming procedures, and most efforts have focused on 3 key approaches: improving MVB/ILV production, preventing degradation, and promoting exocytosis. Exosomes, which originate from early endosomes, undergo gradual maturation to form MVBs that contain numerous ILVs. Ultimately, mature MVBs either fuse with lysosomes for degradation or fuse with the plasma membrane, which results in their release into the extracellular space through exocytosis as exosomes [32, 33].
In this study, we used the Wnt agonist CHIR99021 to prime hucMSCs with the aim of enhancing the production of exosomes as well as their therapeutic efficacy. Our results indicated that pretreatment with CHIR99021 resulted in a significant increase in exosome secretion by hucMSCs, and exosome secretion in this group was approximately 1.5 times greater than that in the control group. This increase in exosome production was achieved without any changes in the characteristics of the hucMSCs or the size of the exosome particles. Furthermore, treatment with CHIR99021 enhanced the intracellular MVB and ILV contents in hucMSCs and promoted the exocytosis of MVBs. This process was partly mediated by SNAP25.
Several previous studies have suggested that activation of the Wnt signalling pathway can facilitate exocytosis in certain cell types, but their numbers are few, and the underlying mechanism is unclear. Zhang et al. discovered that the activation of β-catenin accelerated the assembly of the ESCRT complex and facilitated the exocytosis of 50-nm gold particles [19]. According to reports by Sorrenson et al., β-catenin promoted insulin secretion independently of its transcriptional coactivator TCF7L2; instead, β-catenin somehow facilitated the remodelling of actin and regulated the movement of insulin secretory vesicles towards the plasma membrane [20]. Sándor et al. reported that Wnt pathway activation was coupled with greater EV release in multiple organoid models [21], while Lu et al. suggested that Wnt signalling pathway agonists might exert the opposite effects on EV secretion [22]. In our study, a significant increase in exosome exocytosis was observed in hucMSCs after the Wnt signalling pathway was activated. Further transcriptomics led us to focus on cytosolic exocytosis and SNAP25, which is a crucial component of the SNARE complex. Through the siRNA-mediated knockdown of SNAP25, the CHIR99021-induced increase in exosomal exocytosis was partially abolished, confirming that Wnt agonists might partly rely on SNAP25 to facilitate exocytosis. These results proved that CHIR99021 could be a safe approach for enhancing exocytosis by hucMSCs, thus effectively increasing exosome production. For MVB/ILV production, we observed an increase in the MVB and ILV contents by TEM. Wnt signalling may promote exosome exocytosis and biogenesis; however, the expression levels of the core proteins of the ESCRT complex, namely, ALIX and TSG101, did not significantly change after CHIR99021 treatment. Combined with the decrease in LAMP2-CD63 costaining observed by confocal microscopy, these findings suggested that Wnt agonists increase the MVB and ILV contents by suppressing lysosome-mediated degradation. However, we cannot exclude the possibility that Wnt agonists might regulate the generation of MVBs and ILVs through mechanisms that are independent of ALIX and TSG101 protein synthesis. Other possible pathways include ESCRT-independent pathways, which are facilitated by the generation of ceramide or phosphatidic acid or by lipid raft formation in a Rab31-flotillin-dependent manner [34]. We will explore this topic in our future research.
Next, we explored the effect of CHIR99021 on exosome cargo loading and clinical applications. Proteomic analysis was used to identify the proteins whose levels were altered after Wnt signalling activation, and the proteins that were significantly upregulated in CHIR99021-treated exosomes were associated with pro-healing and cell migration processes. Previous research has demonstrated that activation of the nonclassical Wnt pathway induces the aggregation of IL-6, VEGF, and MMP2 into EVs in melanoma cells. Another study reported that activation of the classical Wnt pathway also promoted the aggregation of miRNAs into exosomes [35]. In our investigation, we observed an increase in the aggregation of TGFβ2, TGFβ3, AGO2, CCAR1, and other pro-healing proteins into exosomes following activation of the classical Wnt pathway. These findings collectively suggested that Wnt signalling has an impact on the sorting of EV contents. However, the specific mechanism underlying this phenomenon is not fully understood and warrants further exploration. For clinical applications, we used a full-thickness cutaneous wound model in diabetic mice, as well as NIH/3T3 fibroblasts, to observe whether the enhanced exosomes had better efficacy. Consistent with previous reports [25, 36, 37], control exosomes from untreated hucMSCs significantly accelerated the wound healing procedure, and our CHIR-Exos were even more effective: compared with control exosomes, CHIR-Exos significantly accelerated the wound healing rate, enhanced local collagen deposition, promoted angiogenesis, and reduced chronic inflammation in diabetic mice. Subsequent in vitro experiments confirmed that the CHIR-Exos promoted wound healing by facilitating cell migration, inhibiting oxidative stress-induced apoptosis, and preventing cell cycle arrest. All of these results suggested that pretreatment with CHIR99021 not only promoted hucMSC-derived Exo production but also improved the therapeutic efficacy of these particles, thus providing a potential approach for clinical application.
In summary, our findings indicated that CHIR99021 could be a safe approach for facilitating exosomal secretion by hucMSCs and augmenting the loading of pro-healing proteins into exosomes to promote diabetic wound healing. However, before we apply these results to biopharmaceutical development and commercial mass production, several challenges need to be addressed. First, although we found that CHIR99021 delayed the passage-induced senescence of hucMSCs, the possibility that exosome cargoes change during continuous subculture still existed. Therefore, exosomal proteomic comparisons are necessary after continuous passaging. Second, the influence of the Wnt signalling pathway on exosomal protein cargo loading and its specific mechanism remain unclear and warrant further exploration. Finally, the precise impact of CHIR99021 on ILV production and its underlying mechanism remain unclear, so further investigations are necessary, particularly regarding the potential involvement of the ESCRT-independent pathway. We hope to address these limitations in our future research.
留言 (0)