CP/CPPS is a complicated and multifaceted disorder with an uncertain origin. An increasing body of research has discovered a potential connection between the gastrointestinal microbiome and various illnesses in humans. Since the brain-gut-bladder axis theory emerged, numerous clinical and animal model studies have validated the correlation between the gut microbiome and bladder symptoms. In recent years, there has been a growing acknowledgement of the connection between gut microbiota and CP/CPPS [19, 20]. But the exact mechanism is not clear. Intestinal flora may influence CP/CPPS via mediating immune inflammatory response, neurotransmitters, androkinin, and direct infection.
Urinary disorders, such as urinary urgency, increased urination frequency, and difficulty initiating or completing voiding, were commonly observed in patients with gastrointestinal disorders [21]. Leue et al. have reported that there is a connection between functional bladder disorders and bowel disease [22]. Considering the frequent occurrence of urinary and colonic dysfunctions together, as well as the possibility of one organ affecting the functioning of the other, it can be concluded that there is an interaction between these two organs. Gut-bladder axis is the most famous hypothesis regarding bladder-gut cross-talk. The gut microbiota play a vital role as an essential mediator in the bidirectional communication between the gastrointestinal tract and the urinary bladder. In these exchanges, inflammasomes, complexes of multiple proteins that can trigger inflammatory reactions, and metabolites produced by the microbiome like short-chain fatty acids (SCFAs), have a significant impact.
SCFAs have become significant microbial byproducts involved in the regulation of immune inflammation and metabolism through interactions with the gut microbiome and host receptors [23]. Within the microenvironment of the proximal colon, three noteworthy SCFAs—acetate, propionate, and butyrate—hold prominence, with concentrations ranging from 50 to 120 mM (mM), showcasing considerable variability within this specified concentration range [24]. They are generated by two primary categories of bacteria, specifically Bacteroidetes and Firmicutes [25]. SCFAs display a versatile capability to influence various aspects of immune cell function, as they possess the ability to modulate gene expression, alter cellular differentiation processes, impact chemotaxis, regulate cellular proliferation, and even induce apoptosis within immune cells, actively participating in the comprehensive process of immune response [26]. According to the findings presented by Du HX et al., the occurrence of gut dysbiosis plays a contributory role in generating an imbalance in the differentiation of Th17 and Treg cells within the context of experimental autoimmune prostatitis (EAP). This imbalance, observed in the differentiation patterns of these immune cells, is intricately linked to the lower levels of propionic acid—a significant SCFA originated from gut microbiota [27]. Chen et al. observed that the oral consumption of glycated whey proteins may induce a prebiotic effect, leading to heightened levels of Allobaculum, Anaerostipes, Bacteroides, Parabacteroides, and Prevotella, while concurrently lowering the levels of Adlercreutzia and Roseburia at the genus level. Consequently, this led to a decrease in the immune inflammatory response of mice in the EAP model [28].
Interestingly, 4 GM taxa, including family Christensenellaceae, genus Eisenbergiella, genus Terrisporobacter, and genus hungatella of our results, all belong to the phylum Firmicutes. In the healthy population, there was a greater presence of Christensenellaceae, which showed an inverse correlation with inflammation [29]. Kropp C et al. reported that Christensenellaceae minuta has the ability to inhibit intestinal harm, decrease inflammation in the colon, and facilitate the healing of the mucosal layer in both in vitro and in vivo experiments [30]. Similarly, Relizani K et al. presented a comprehensive screening procedure that integrates in vitro and in vivo tests to systematically choose a viable strain of Christensenellaceae minuta with potent immunomodulatory characteristics. This indicates that Christensenella minuta has the potential to be utilized as a future biotherapy for Crohn’s disease [31]. The relationship between Genus Eisenbergiella and eubiosis is likely due to its ability to generate significant metabolic products such as butyrate, acetate, lactate, and succinate, which have a nourishing impact on the mucosa[32]. Moreover, the genus Eisenbergiella might be strongly associated with the observed reduction in inflammation of intestine among mice with ulcerative colitis [33]. In accordance with Eisenbergiella, the genus Terrisporobacter was found to have significantly linked to SCFAs and oxidative stress in a study involving animals[34]. Qiao J et al. reported that rapeseed bee pollen can inhibit pathogenic bacteria and enhance probiotics, particularly in the Firmicutes-to-Bacteroidetes (F/B) ratio to alleviate chronic non-bacteria prostatitis [19], which is basically consistent with our findings. However, to the best of our knowledge, there have been no prior examinations exploring the potential link between the genera Eisenbergiella/Terrisporobacter and CP/CPPS.
CP/CPPS may be also related to abnormal release of neurotransmitters. The main neurotransmitters involved in the occurrence and development of CP/CPPS are “5⁃hydroxytryptamine(5⁃HT), noradrenaline (NE), and dopamine (DA).” They have a significant impact on this condition. Statistics show that about 40% of CP/CPPS men suffer from premature ejaculation [35]. Moreover, 78% of CP/CPPS patients exhibit psychological distress, including anxiety and depression [36]. The concentrations of 5⁃HT and various other neurotransmitters in both peripheral blood and the brain may be subject to influence by the gut microbiota. Yano et al. showed that the 5⁃HT level in germ-free mice was lower than that in healthy mice. When normal intestinal flora was transplanted into germ-free mice, it was possible to restore it to normal levels [37]. Du et al. reported that the EAP mice showed obvious depression-like behavior and the composition of intestinal bacteria such as Dantamoesophagota, Ruminococcus, and Bacteroidetes was significantly different from that in normal mice. Fecal bacteria from EAP mice transplanted into pseudogerm-free mice treated with antibiotics could aggravate the depression-like behavior of the host [38]. Disrupted neurotransmitter release can impact the functioning of the nervous system, leading to symptoms of depression and sexual dysfunction in individuals with CP/CPPS. 5⁃HT reuptake inhibitors can be used for targeted treatment [39].
Besides, gut microbiota correlate with androgen levels, which regulates prostate. Sufficient androgens can promote prostate growth and development, maintain secretion and differentiation function of prostate. Androgen imbalance can cause CP, the mechanism may be caused by the decrease of androgen content level, which has the effect of inhibiting humoral immunity, and when its content is reduced, it can cause autoimmune reaction. In addition, androgen can promote prostate autophagy, while high levels of estrogen inhibiting autophagy [40, 41]. Once androgen level decreased and the balance of male and female hormones disturbed, prostate cells may be damaged due to insufficient autophagy, inducing prostate inflammation. Poutahidis et al. found that the serum testosterone level was significantly increased after feeding Lactobacillus reuteri to mice [42]. Konkol Y et al. induced nonbacterial chronic prostate inflammation (CPI) in the Wistar rat strain for 18 weeks with subcutaneous testosterone and 17β-estradiol (E2) hormone pellets. A decrease in Bacteroides uniformis, Lactobacillus, and Lachnospiraceae levels was observed in rats suffering from CPI. In the fecal samples from those rats, SCFA butyric-, valeric-, and caproic-acid concentrations were also decreased [20]. These results and theories are similar to our findings. Liu et al. demonstrated that poria cocos polysaccharides and finasteride had the ability to alter the composition of intestinal flora in a rat model of CP/CPPS. This led to a decrease in the levels of pro-inflammatory cytokines (TNF⁃α, IL⁃2, and IL⁃8). Simultaneously, there was a reduction in androgens (dihydrotestosterone and testosterone), resulting in an improvement in both prostatic inflammation and histological damage, which is suggested that CP/CPPS can be treated by regulating the gut microbiota and androgen level [43].
Finally, gut microbiota may affect CP/CPPS through direct infection. Although it has not been confirmed that there is a direct pathway between the rectum and the prostate to explain how gut microbiota infuse the urogenital tract. The two organs are anatomically adjacent, and there is the possibility of “direct infiltration” [44].
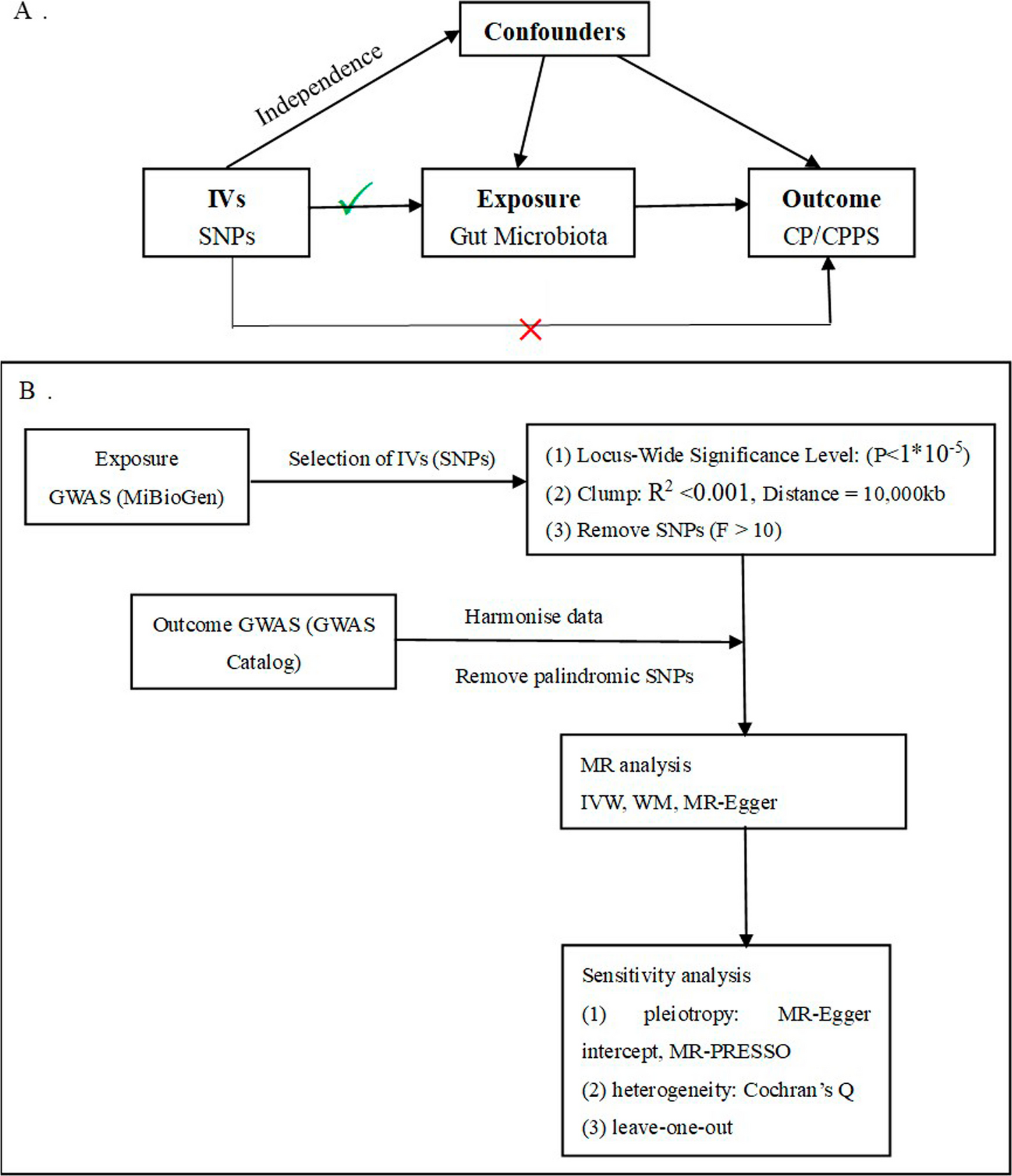
In the current study, we utilized the summary data of gut microbiota from the MiBioGen Consortium’s extensive GWAS meta-analysis and the GWAS Catalog's summary data of CP/CPPS to investigate the causal link. Mendelian randomization and sensitivity analysis were performed to assess the causal relationship between gut microbiota and CP/CPPS. Protective effects against CP/CPPS were observed in various genera of gut microbiota, including ChristensenellaceaeR.7group, Eisenbergiella, Hungatella, Terrisporobacter (OR < 1), whereas the family Prevotellaceae (OR > 1) had the opposite effect.
According to recent researches, rheumatoid arthritis [45], periodontitis [46], and intestinal and vaginal dysbiosis [47,48,49] are linked to the higher presence of Prevotellaceae family members in various microbial ecosystems. Prevotella colonization may lead to metabolic alterations in the microbiome, resulting in decreased IL-18 synthesis. This, in turn, exacerbates intestinal inflammation and contributes to the development of systemic autoimmunity. Regrettably, given the intricacy of gut microbiota, there is indeed a lack of agreement between our findings and the current supporting data. For example, Shoskes DA et al. used 16S rRNA sequencing, discovering reduced gut microbiota richness and diversity in CPPS patients compared to controls, along with markedly diminished levels of Prevotella [50]. Hence, additional research is required to authenticate these connections.
Our research brings distinct strengths. Firstly, through a two-sample MR analysis, we amplified the influence of gut microbiota on CP/CPPS causality. Secondly, the strict quality control procedures and robust MR methods applied ensure the reliability and stability of the causal estimates. Thirdly, the identification of potential causal links through the IVW method serves as a guiding beacon for further exploration into specific gut bacteria. This enlightens the investigation of their roles in shaping the intricate landscape of CP/CPPS development.
However, this task also possesses compelling constraints. While meeting the MR assumptions by ensuring the strong correlation of instrumental variables with gut microbiota taxa, our study remains vulnerable to the absence of a guarantee against weak instrumental bias. Firstly, the relatively modest sample size of CP/CPPS may cause some bias in the selection process of IVs, and there may be weak instrumental variables, which may reduce the reliability of the results, impeding the full scope of our research insights. Secondly, the reverse MR analysis was impeded by a limited number of IVs, preventing the establishment of a potential reciprocal causal connection between CP/CPPS and gut microbiota. Thirdly, although MR techniques can offer fresh perspectives on the causal relationships between exposure characteristics and outcome characteristics, the accuracy of estimating the strength of these associations may be limited. Therefore, further investigation is necessary to validate the results. Fourthly, it is crucial to recognize that the predominant participants in this GWAS hail from European ancestry. This demographic specificity raises a noteworthy consideration—the outcomes of this study may not be universally applicable to ethnically diverse populations. Finally, due to the lack of specific country, region, age, and other information, stratification analysis cannot be carried out, which may cause bias to research results. These limitations currently present hurdles in definitively grounding the cause–effect correlation between gut microbiota and CP/CPPS, urging further research endeavors that encompass a broader spectrum of ethnic backgrounds for comprehensive insights. In the future, if a GWAS dataset of CP/CPPS is publicly released with a larger number of participants and SNPs, or from other ethnic groups, it is hoped that additional researches can confirm these connections.

留言 (0)