Identifying germline mutations in high and moderate-risk BC genes is of paramount importance for establishing genetic screening programs that facilitate early diagnosis and development of national public health policies. Implementation of genomic analysis through NGS and incorporation of noBRCA genes has proven to be an adequate strategy to increase sensibility regarding recurrent mutation analysis restricted only to BRCA1/2 genes [12, 13].
Globally, germline mutation cancer prevalence, can be estimated from hereditary, familial, or unselected BC cases. European, North American, and Asian populations have been the primary focus to obtain this data.
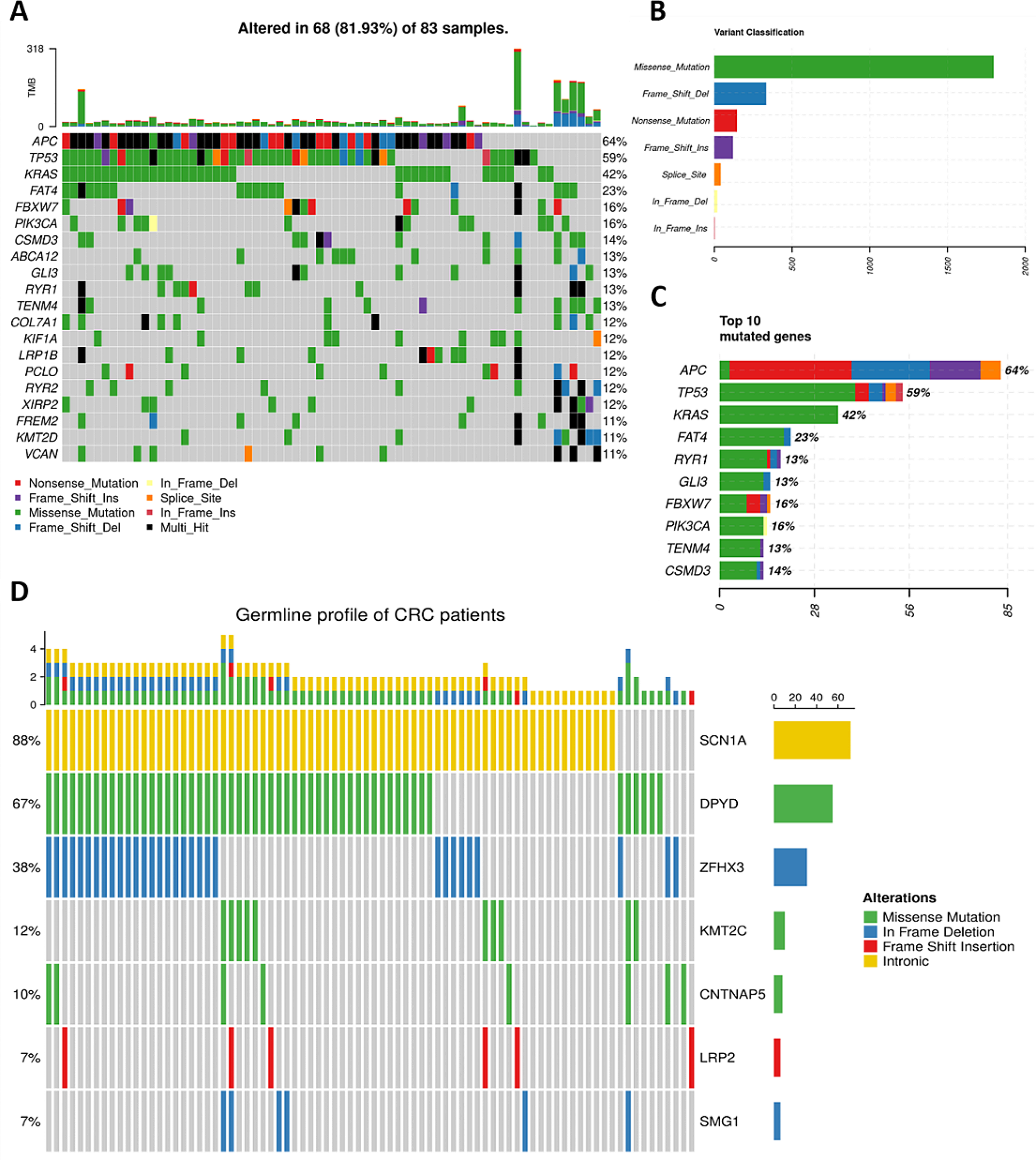
To our knowledge, this is the first report on the prevalence of mutations in the top 10 clinically impactful genes, identified by WES in 400 women with unselected BC from various regions of Colombia.
We evaluated NCCN criteria [10] in the women studied. Significantly, 20.8% of them with a P/LP variant did not fulfill those criteria. This finding demonstrates that molecular testing should be considered in all women with BC regardless of the age of diagnosis, molecular subtype, and personal or family history of cancer.
Our findings determined that 6% of the Colombian women with unselected BC had germline mutations in seven of the genes studied, being BRCA2 the gene with the highest frequency of variants and women affected, followed by ATM, BRCA1, PALB2, CHEK2, BARD1 and RAD51D genes. No P/LP variants were detected in CDH1, RAD51C, and TP53 genes. BC prevalence of germline mutations and their frequency in cancer risk genes, varies thoroughly depending on the selection criteria of the population studied.
Interestingly, to date at least, 41 articles have been described that analyze genes related to BC in the Latin American population. This includes a diversity of patients from Argentina, Brazil, Chile, Guatemala, Colombia, Peru, Puerto Rico, and Mexico, covering 40% of the countries considered in the region through genetic analysis (Supplementary Table 1, and the references therein). These studies have examined approximately 51,000 Latin American patients, which have provided insights into the frequencies of molecular variants of interest in the analyzed genes (BRCA1/2 and noBRCA) (Supplementary Table 1 and the references therein). Concerning the mutational spectrum exhibited by the BRCA1/2 genes, a range from 10.1% to 37.2% has been noted across the populations. This variation is estimated to be strongly linked to the migration history of the Latin American populations, including the overlap of some mutations determined by shared events and exchanges that characterize the migration history of each geographical region [14]. Additionally, within the same population, such as Brazil, there is high variability in the mutation frequencies of the BRCA genes (10.1% vs 22.4%), supporting the observation that the genetic background of Latin American populations results from events leading to unique population structures within and between countries [14,15,16]. Specifically, the highest frequencies for the BRCA genes reported in the Latin American population are described in patients with breast and ovarian cancer from Afro-Colombian families, in whom 33.3% of pathogenic variants were identified [17], demonstrating the impact of patient selection criteria on the variability of reported data.
Unlike our study, most studies reported in Latin America have involved patients with hereditary BC, in whom the representation of pathogenic variants in the BRCA genes is substantially higher than in cases of unselected BC. For this latter group, frequencies between 1.2 and 14.5% have been reported (with eight studies in Latin America) [6, 18,19,20,21,22,23,24], which is consistent with the findings identified in the present study (Table 2 and supplementary Table 1). The analysis of unselected populations has been recommended to avoid the overestimation of the true prevalence of germline cancer-related P/LP variants in the general population [25].
In Colombia, previous reports described mutations in BRCA1/2 genes focused on hereditary/familial cases [17, 21, 26, 27]. Even though, few studies analyzed mutation prevalence in BRCA1/2 genes from unselected BC patients, finding that their frequency ranges from 0.4 to 3.3% [20, 21], which is concordant with our results, since the frequency of women with mutations in BRCA1/2 is 3.25%.
Beyond BRCA genes in Latin America NGS multigene analysis has been conducted in 78% of studies, including the current study, which has enabled the identification of P/LP variants in moderate and low cancer-risk genes, potentially actionable [28]. Our study demonstrated that while 52.3% of the P/LP variants were associated with BRCA1/2 genes, nearly 50% of the women had mutations in noBRCA genes. These findings are similar to those reported in unselected BC populations from countries such as Argentina and Guatemala, where the contribution of noBRCA genes was described as 4.7% and 3.2%, respectively [6, 19]. Similar to studies concerning hereditary BC cases, the frequency of P/LP mutations in noBRCA genes constitutes a significant proportion (Supplementary Table 1). Paixão et al. (2022) found P/LP variants from 9.6% (BRCA1/2) to 25.2% (noBRCA) analyzing 321 Brazilian patients with a panel of 94 genes [7]. Additionally, Cock-Rada and colleagues assessed 25 cancer susceptibility genes in 85 women from Medellin, who met the criteria for HBOC molecular testing; this study identified mutations in six genes: BRCA2, BRCA1, PALB2, ATM, MSH2, and PMS2 [29]. All these findings describe germline mutation profiles which, like our results, demonstrate the contribution to the genetic variability in BC of genes such as ATM, PALB2, and CHEK2, and should be taken into consideration. This finding is consistent with reports from other Latin American populations, where mutations in PALB2 or RAD51C explain a significant proportion of cases. The present results, along with others previously published, demonstrate that the analysis of genes other than BRCA1/2 increases the detection rate of P/LP variants, which maximizes the identification of germline variants in patients with hereditary and unselected BC.
In our study, we identified recurrent mutations in 1.75% of the population analyzed, indicating that most P/LP variants are private. This finding is consistent with previous reports in other LATAM populations, where the recurrence of mutations is low [13]. Three recurrent variants were identified: two in the BRCA2 gene (c.2808_2811delACAA, p.Ala938Profs*21 and c.1763_1766delATAA, p.Asn588Serfs*25), and one in the ATM gene (c.5496 + 2_5496 + 5delTAAG). Two women carrying recurrent mutations in BRCA2 (c.1763_1766delATAA, p.Asn588Serfs*25) and ATM (c.5496 + 2_5496 + 5delTAAG) genes had the diagnosis of other types of cancer that is, lymphoma and, thyroid and gastric, respectively. Co-occurrence between BC and other types of cancer has been pinpointed in the literature [30, 31]. Specifically, P/LP variants in the ATM gene are associated with gastric and thyroid cancers, and risk estimates have also been described; for gastric cancer, several studies associated ATM mutations with OR (odds ratio) ranging from 2.97 to 4.74 [32,33,34]. Recently, the association between H. pylori infection and germline P variants in genes as BRCA1, BRCA2, ATM, and PALB2, has been described; people with H. pylori infection and germline mutations in those genes have a higher gastric cancer cumulative risk at 85 years of 45.5% (95% CI, 20.7 to 62.6); in contrast, the risk in people with H. pylori infection alone is 14.4% (95% CI, 12.2 to 16.6) [35]. Thyroid cancer (TC) has also been associated with the presence of germline mutations in BRCA2 and ATM genes [36, 37]. Interestingly, a published study showed an increased oncogenic SNPs burden in cases with co-occurrence of BC and TC. In patients with double cancers, germline variants were found in PALB2, BRCA1, BRCA2, ATM, and CHEK2 genes, which are known risk genes associated with BC [38].
Recurrent variants could also be considered founder mutations. The prevalence of founder mutations has been extensively documented for the BRCA1 and BRCA2 genes. These pathogenic variants represent the majority of observed mutations in specific populations and have been confirmed as true founders through analysis of common ancestral haplotypes [39]. In our population of study three Colombian founder mutations, previously described [21], were identified, one in BRCA1 c.5123C > A (A1708E), and two in BRCA2 c.1763_1766delATAA (1991del4) and c.2808_2811delACAA (3034del4).
Identification of recurrent pathogenic variants in the ATM gene is of importance, as previous studies have demonstrated that women carrying mutations in this gene have a significantly increased risk of developing BC with a risk similar to that conferred by germline mutations in the BRCA2 gene [40]. Interestingly, the allelic frequency of the ATM variant c.5496 + 2_5496 + 5delTAAG was 0.375%, although it has not been previously reported in the population database gnomAD, the variant has been identified in cases related to ataxia-telangiectasia syndrome, familial breast cancer, and hereditary cancer predisposition syndrome. These findings are not supported by population-based studies but have been submitted by molecular diagnostic centers such as Color Diagnostics (2019), Fulgent Genetics (2021), Baylor Genetics (2022), Invitae (2022), Ambry Genetics (2023), and Myriad Genetics (2024) (https://www.ncbi.nlm.nih.gov/clinvar/variation/VCV000565770.15 (accessed May 7, 2024)). In all instances, the variant has been determined to be germline. However, due to the unknown origin, the number of affected individuals, or the lack of familial segregation analysis, we cannot make comparisons with the data from the current study. It is noteworthy to date, this variant has been attributed to a significant impact on RNA splicing, although this has not been experimentally proven, hence its classification according to ACMG criteria is likely pathogenic. Functional validation of this recurrent variant demonstrated an exon skipping, leading to a predicted deletion of 59 amino acids located in the Pincer domain of the ATM protein [41]. The splicing process is an event that most eukaryotes genes go through and is regulated by RNA-Binding Proteins (RBPs), cis-regulatory elements, and trans-acting factors [42]. Alternative splicing is dysregulated in cancerous cells in comparison with healthy cells, and carcinogenesis has been associated with alterations in direct and indirect regulators, leading to altered splicing profiles [43]. In the present study, the minigene assay resulted in an exon skipping, caused by a cis-regulatory element (c.5496 + 2_5496 + 5delTAAG) on the ATM gene. This molecular finding added to the absence of this mutation in the gnomAD database, supports the pathogenic effect of the mutation in the function of the ATM protein and the possible role in BC development. Dysregulation of alternative splicing in cancer has made it a therapeutic target and several therapeutic strategies are currently under study; that is, targeting RNA splicing factors, splicing factors regulated by blocking kinases, and antiRNA molecules [44].
Although 58.3% of women with a mutation had several relatives with various types of cancer, segregation analysis was performed in some families with index cases having P/LP variants in BRCA2, ATM, and PALB2 genes. Interestingly, segregation of P/LP variant and phenotype was observed in two families tested for BRCA2 mutations (Fig. 1). Index cases of these families had an earlier age of onset compared with their relatives who suffered BC as well, suggesting anticipation phenomena. This finding may be associated with the greater penetrance of the BRCA1/2 genes, compared to other genes with moderate penetrance such as ATM, but some authors have proposed the interference of non-genetic factors as an explanation for this anticipation [45, 46].
Germline pathogenic small indels and LGRs contribute to the development of breast and ovarian cancers [47]. Ratios of BRCA1/2 LGRs germline mutations are population dependent [22, 48,49,50,51,52,53,54,55,56]. To our knowledge, in Colombia, BRCA1/2 LGRs have been tested in two studies. Vargas and colleagues tested 60 Afro-Colombian families with HBOC, they did not find LGRs in that population [17]. Torres and colleagues tested 221 breast/ovarian cancer families, finding a LGR in the BRCA2 (ex1-14del) gene in two unrelated patients (0,9%) [21]. Considering the three Colombian cohorts of patients assessed for BRCA1/2 LGRs (Vargas et al., Torres et al., and ours), the prevalence of this type of rearrangement in BRCA1/2 genes would be 0,3% (2/681). Pondering the frequencies described previously, LGRs prevalence in BRCA1/2 genes is low in Colombian BC patients, regardless of hereditary or family history.
This study has some limitations. The germline variants analyzed are rare and although they are located in high and moderate-risks genes, common SNPs also contribute to the development of BC. LGRs were only studied in BRCA1/2, although this type of rearrangement has been found in genes including CHEK2 and ATM, in BC patients [57].
In conclusion, molecular analysis via WES enabled the establishment of the genomic profile of P/LP variants in ten clinically significant genes related to BC risk in the analyzed population. Additionally, this investigation was conducted in a population of women with unselected BC, which has been less addressed in the global literature compared to the vast amount of research conducted on individuals with hereditary cancer. Based on the information described and our study results, the germline mutation profile exhibits variation in genes and frequencies, contingent upon the region and characteristics of the population assessed. This underscores the importance of conducting population-based studies and determining the prevalence of clinically impactful genes. Such efforts can aid in the identification of mutations and facilitate the implementation of national genetic analysis policies, genetic counseling, and early detection strategies. Our study also highlights the utility of WES as an appropriate method for identifying germline variants located in coding and exon–intron boundary regions of genes that are clinically relevant in BC. WES analysis has the potential to detect rare, novel, and infrequently studied P/LP variants, including intronic mutations.

留言 (0)