
記住我






To obtain inducible HO-1 expression in SCs of dystrophic mice, we generated double-transgenic mice on mdx background (Fig. 1A). In brief, ROSA26LSL-HO1 mice with human HMOX1 cDNA downstream of a floxed stop codon cloned into the endogenous ROSA26 locus were generated previously [18]. To excise the stop codon and enable HMOX1 transgene expression, we introduced Cre recombinase in Pax7-CreERT2 mice with an internal ribosome entry site (IRES)-CreERT2 fusion construct inserted downstream of the stop codon of Pax7 gene. Pax7, a SC marker, is expressed and functional, while Cre-ERT2 activity is inducible by tamoxifen (TX), a selective estrogen receptor regulator, that allows Cre nuclear localization. Dystrophic background was introduced by mdx mice breeding with Rosa26LSL-HO1 and Pax7-CreERT2 mice (Fig. 1A).
Obtained double-transgenic mouse model (mdx;HMOX1Pax7Ind) allows TX-inducible, Cre-mediated recombination at the LoxP sites, and HMOX1 expression in Pax7 positive cells. In parallel, TX-treated mdx mice served as control of potential side effects of TX.
The body weight of untreated animals (before experiment) was higher in mdx;HMOX1Pax7Ind than in mdx mice (Fig. 1C). However, in both genotypes neither TX- nor exercise-dependent effect on body weight was detected as measured before and two weeks after the last TX injection (Suppl. Figure 1A). In addition the total weight of hind limb skeletal muscles (caput gastrocnemius, soleus, tibialis anterior, EBL, quadriceps, adductor) collected from one limb measured at the end of experiment was comparable between groups of both genotypes (Suppl. Figure 1B).
TX induces HMOX1 expression in skeletal muscles of transgenic mdx;HMOX1 Pax7Ind miceTransgenic and control mice aged 13–16 weeks were subjected to the acute physical exercise protocol with high-speed treadmill (12 m/min, 45 min) and downhill running [19] to worsen skeletal muscle phenotype and reveal immediate effects of HO-1 on muscle pathology and SC activation. Mdx;HMOX1Pax7Ind and mdx mice were given 5-day TX injections and upon 7-day waiting subjected to exercise protocol (Fig. 1B).
Comparable mRNA level of murine Hmox1 was detected in mdx mice and transgenic mdx;HMOX1Pax7Ind mice both in caput gastrocnemius (GAS) (Fig. 1D) and tibialis anterior (TA) (Fig. 1E). TX administration induced the expression of human HMOX1 (over oil group) specifically in skeletal muscles of transgenic mice, GAS (Fig. 1F) and TA (Fig. 1G). Nonetheless, HMOX1 expression in oil group of both transgenic muscles was higher than in oil-treated mdx counterparts suggesting either the leakage of the Cre-LoxP system and TX-independent action of Cre, or HMOX1 DNA detection (Fig. 1F,G).
In mdx mice, murine Hmox1 was diminished in skeletal muscles on day 3 upon treadmill exercise, with TX exacerbating this decline (Fig. 1H) and opposite trends seen in mdx;HMOX1Pax7Ind mice (Fig. 1I). One may consider the regulation of Hmox1 by human HO-1 activity in transgenic mice or strain-dependent pattern of Hmox1 regulation.
No HMOX1 expression was detected in mdx mice (Fig. 1J). The induction of human HMOX1 expression after TX treatment was detected in skeletal muscles of untreated transgenic mice and further increased upon exercise (Fig. 1K). This might be related to enhanced activity of ROSA promoter after exercise but requires more in-depth analysis.
TX increases HO-1 level in SCs of transgenic mdx;HMOX1 Pax7Ind miceHO-1 protein was detected in ~ 20% of all nucleated cells (Suppl. Figure 2A,B) and at similar proportion in white blood cells (WBCs, CD45+CD31−) (Fig. 2A) in hind limb skeletal muscles (GAS, TA, soleus, EBL, quadriceps, adductor) of transgenic as well as control mdx mice. The number of HO-1+ cells in WBC population increased further by physical exercise in mdx mice (Fig. 2A). Interestingly, the majority (~ 90%) of CD45−CD31−Sca-1+, that may comprise side population SCs [20] or myoblasts [21], of both strains expressed HO-1 (Fig. 2B, Suppl. Figure 2A). However, in the transgenic animals we detected exercise-specific drop of CD45−CD31−Sca-1+ population expressing HO-1, not seen in mdx counterparts (Fig. 2B).
Fig. 2
HMOX1 expression in induced by TX in SCs of mdx;HMOX1Pax7Ind mice. Quantitative analysis of the percentage of HO-1 positive cells among (A) CD45 + CD31- (WBCs), (B) CD45-CD31-Sca-1 + cells, and (C) representative plots and (D) quantitative analysis of the percentage of HO-1 positive cells among SCs in mdx and mdx;HMOX1Pax7Ind hind limb skeletal muscles. Flow cytometry (n = 5–8). Data is presented as mean± SEM. ## p < 0.01, ### p < 0.001—Two-way ANOVA interaction; *** p < 0.001 by Two-way ANOVA with Tukey’s post hoc test. (F) HO-1 is expressed and colocalizes with Pax7 expression in TX-treated mdx;HMOX1Pax7Ind but not in mdx mice. Immunofluorescence of Pax7 (green), HO-1 (red) and nuclei (DAPI, blue). Scale bar: 5 μm
The specific induction of HO-1 was detected in SCs (CD45−CD31−Sca-1−α7integrin+) (Suppl. Figure 2A), upon TX administration both in untrained and exercised transgenic mice (vs. oil treatment and mdx counterparts) (Fig. 2C,D). As in the case of HMOX1 mRNA level in GAS and TA (Fig. 1F,G), the HO-1 protein in SCs could be already detected in oil-treated mdx;HMOX1Pax7Ind mice (vs. mdx) (Fig. 2D). Finally, HO-1 protein was detected in Pax7-positive cells in TX-treated mdx;HMOX1Pax7Ind but not in mdx mice (Fig. 2E) verifying SC-targeted HO-1 induction in mdx;HMOX1Pax7Ind skeletal muscles.
HO-1 induction decreases SC pool in mdx;HMOX1 Pax7Ind miceThe high-speed treadmill exercise regimen decreased SC pool in control, oil-treated mice of both HO-1 transgenic and control mice (Fig. 3A). HO-1 induction caused a drop in SC pool, decreasing the percentage of SCs in mdx;HMOX1Pax7Ind mice (Fig. 3A) to the comparable extent as the physical training (Fig. 3A).
Fig. 3
The effect of HO-1 induction on SC pool, activation and proliferation. A HO-1 decreases SC pool in hind limb muscles of mdx;HMOX1Pax7Ind mice. Quantitative analysis of the percentage of SCs in muscles of mdx;HMOX1Pax7Ind and mdx mice. Flow cytometry (n = 5–8). B SC activation (based on the loss of CD34) and proliferation (Ki67 +) of (C) SCs and (D) CD34- SC subpopulation in hind limb skeletal muscles of mdx and mdx;HMOX1Pax7Ind mice. Flow cytometry (n = 5–8). Data is presented as mean± SEM. # p < 0.05, ## p < 0.01, ### p < 0.001—Two-way ANOVA variation; * p < 0.05, ** p < 0.01 by Two-way ANOVA with Tukey’s post hoc test
Studies have shown that high intensity physical exercise that induce muscle insult is associated with SC activation and increase in numbers during the acute phases of injury [7,8,9]. In HO-1 transgenic (mdx;HMOX1Pax7Ind) and control mdx mice, SC activation associated with the loss of CD34, a marker of quiescent SCs [22], increased in response to physical exercise in a comparable manner (Fig. 3B). CD34− SCs were also more frequent in both genotypes upon TX administration suggesting HO-1-independent stimulation of SC activity (Fig. 3B).
No HO-1 specific effect was detected in populations defined upstream SC gating: CD45−CD31− (Suppl. Figure 3A), CD45+CD31− (WBCs, Suppl. Figure 3B), CD45−CD31+ (endothelial cells, ECs, Suppl. Figure 3C), CD45−CD31−Sca-1− (Suppl. Figure 3D) or CD45−CD31−Sca-1+. However, TX treatment caused a decrease of the percentage of CD45−CD31−Sca-1+ in mdx mice, the effect reversed by physical exercise (Suppl. Figure 3E).
Upon physical exercise, the percentage of proliferating SCs (Fig. 3C) and CD34− SC subpopulation (Fig. 3D) decreased in control mdx mice, an effect that was reversed in mdx;HMOX1Pax7Ind mice. Similar pattern was detected in oil-treated and TX-treated mdx;HMOX1Pax7Ind mice that could be related to the leakage of the Cre-LoxP system (Fig. 3C,D, compare with Figs. 1F,G, 2D).
In skeletal muscles of control mdx mice, we observed exercise-dependent decrease in murine Hmox1 expression that correlated with a drop in SC proliferation (Figs. 1H, 3C,D). In contrast, exercise induced HO-1 mRNA level in transgenic mice and also increased SC proliferation (Figs. 1I, K, 3C, D). On the other hand, no respective changes in HO-1 level was visible in SCs (Fig. 2D).
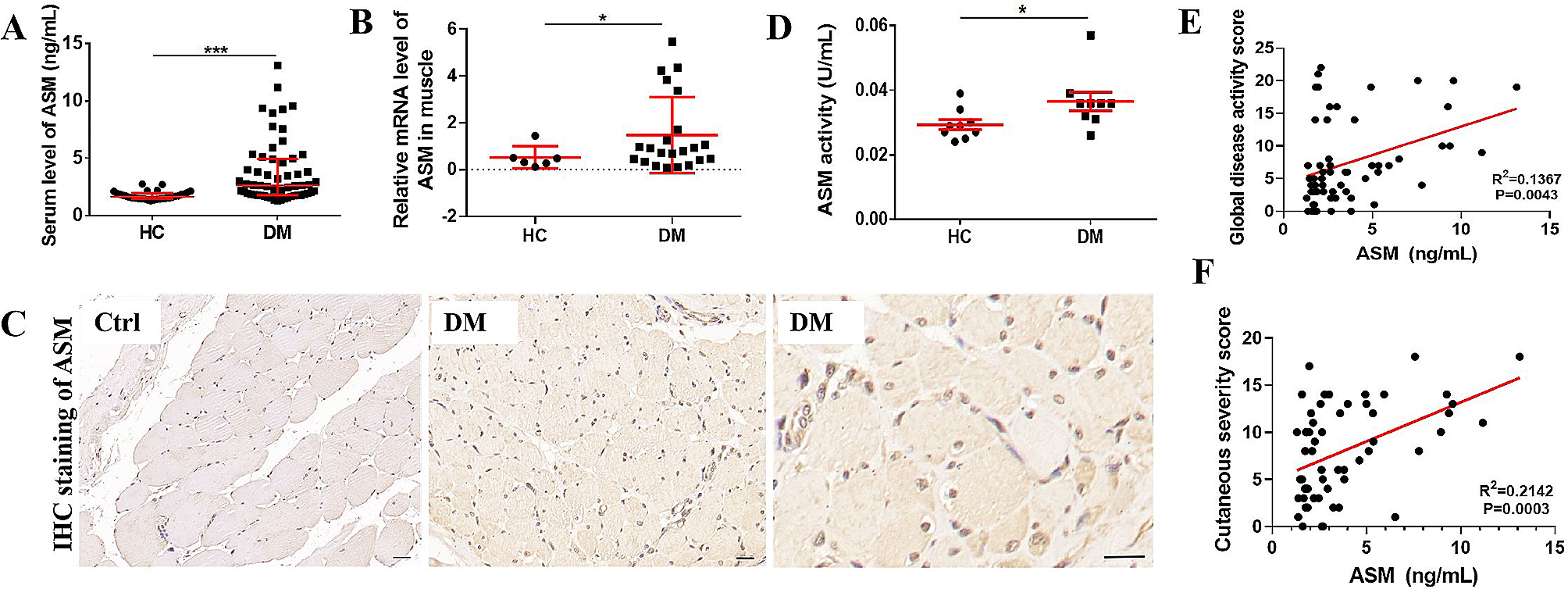
SC-targeted HMOX1 expression modulates skeletal muscle inflammation in dystrophic miceMuscle-derived circulating CK and LDH serve as DMD biomarkers reflecting membrane damage, necrosis and extracellular leakage [23, 24]. Mdx;HMOX1Pax7Ind mice showed lower serum level of muscle damage markers upon physical exercise in comparison to mdx mice (Fig. 4A,B).
Fig. 4
mdx;HMOX1Pax7Ind mice show lower serum level of muscle damage markers upon physical exercise. A LDH and (B) CK activity in serum of mdx and mdx;Pax7Ind mice. Colorimetric evaluation (n = 5–9). Data is presented as mean± SEM. # p < 0.05—Two-way ANOVA variation; * p < 0.05 by Two-way ANOVA with Tukey’s post hoc test
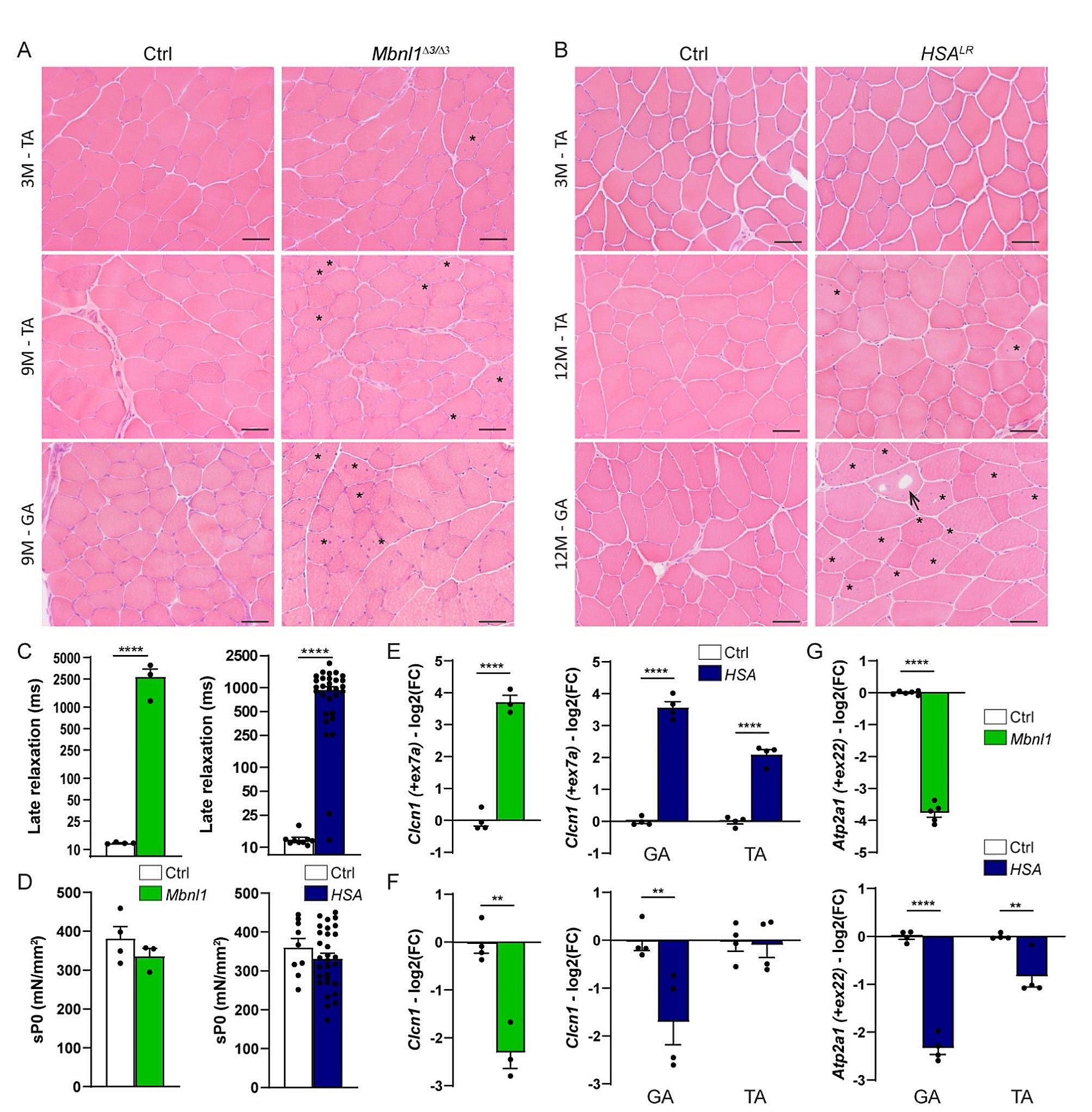
To further outline the effect of HO-1 in SCs we characterized a dystrophic phenotype and the extent of skeletal muscle damage in mdx;HMOX1Pax7Ind mice (vs. mdx counterparts). The percentage of WBCs (Suppl. Figure 3B) as well as ECs (Suppl. Figure 3C) in hind limb muscles assessed by flow cytometry was not changed by SC-specific HO-1 induction neither in untrained nor in exercise groups and was comparable to mdx counterparts (Suppl. Figure 3B,C). The histological analysis of TA, GAS and DIA muscles of mdx and mdx;HMOX1Pax7Ind mice did not reveal significant effects of HO-1 on inflammatory response (Fig. 5A-D). However, there was an increase in inflammatory cell infiltration after exercise in TA of mdx mice, but not in HO-1 transgenic mice (Fig. 5B). Also, the basal inflammation extent was moderately higher in GAS of mdx mice than in HO-1 transgenic mice (Fig. 5A,C). In addition, HO-1 transgenic mice revealed anti-inflammatory cytokine profile. The level of antinflammatory interleukin 10 (IL-10) known to limit T cell activation and production of proinflammatory cytokines [25] increased in mdx;HMOX1Pax7Ind mice (vs. mdx counterparts) both in response to exercise and TX administration (Fig. 5E). In parallel, TX increased IL-10 level in untrained mdx;HMOX1Pax7Ind mice, while decreased run-induced IL-10 expression suggesting more complex inflammatory effect (Fig. 5E).
Fig. 5
HMOX1 induction in SCs modulates skeletal muscle inflammation in dystrophic mice. A-E Inflammation assessment in tibialis anterior (TA), diaphragmae (DIA) and gastrocnemius (GAS) tissue sections. (A) Representative images of leukocyte infiltration/necrosis in skeletal muscles. Scale bar: 100 μm. B-D Semiquantitative analysis in (B) TA, (C) GAS and (D) DIA (n = 5–9). H&E staining. Relative expression of Il10 in TA. qPCR (n = 3–7). Data is presented as mean± SEM. # p < 0.05, ## p < 0.05—Two-way ANOVA variation; ** p < 0.01 by Two-way ANOVA with Tukey’s post hoc test
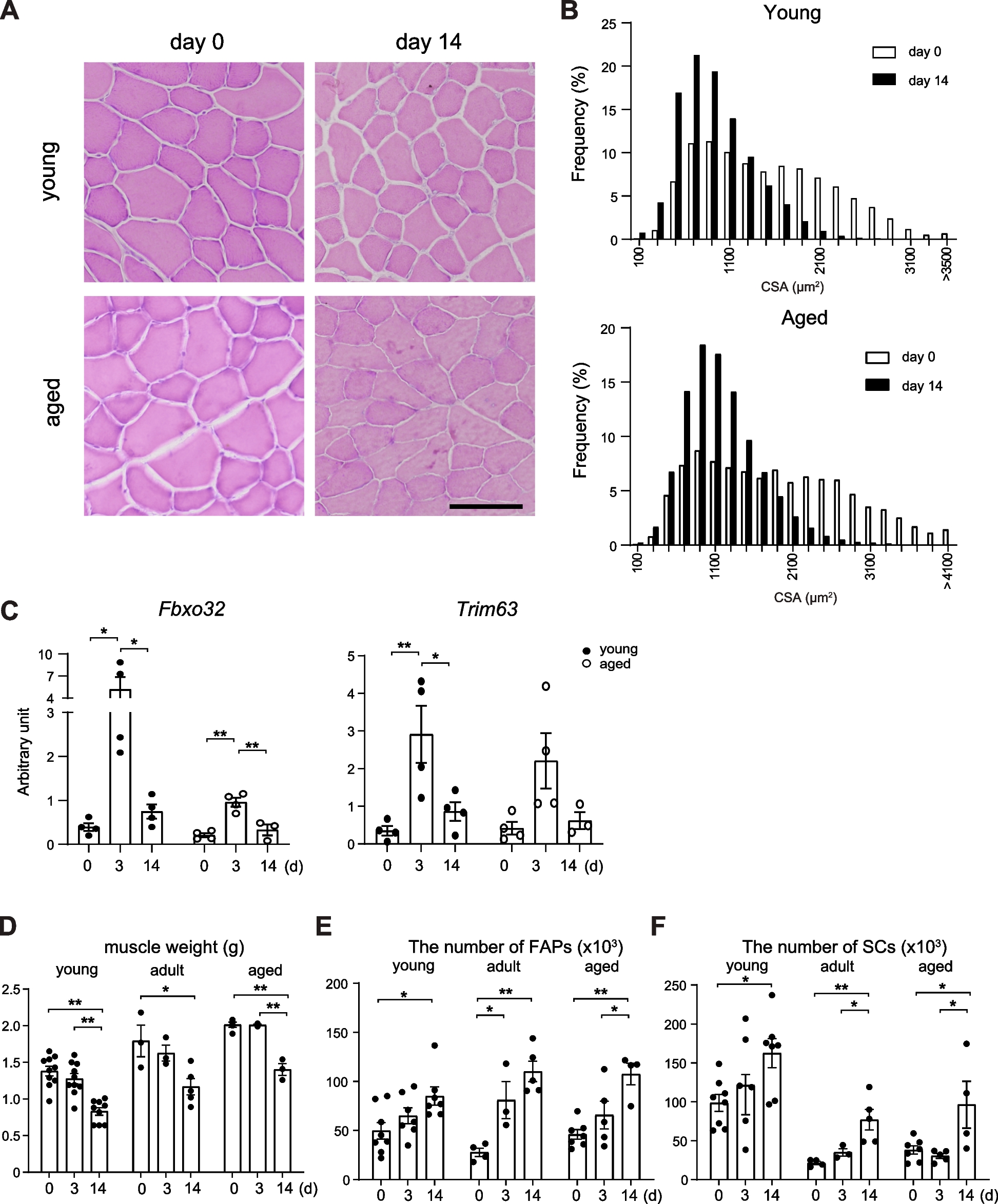
SC-specific HMOX1 induction attenuates skeletal muscle necrotic events upon physical exerciseThe analysis of skeletal muscles revealed HO-1 dependent anti-necrotic action upon physical exercise in tibialis of mdx;HMOX1Pax7Ind and opposite pattern seen in TX-treated mdx mice, as evidenced by assessment of IgM and IgG accumulation (Fig. 6A-E). In untreated mdx mice necrotic events decreased in TA after TX treatment while rose back to control values upon physical exercise, in contrast to HO-1 transgenic mice, in which the IgG/IgM accumulation decreased significantly after exercise (Fig. 6A-C). The reverse effect of TX in mdx;HMOX1Pax7Ind and mdx mice in TA (Fig. 6A-C) resembled the pattern of inflammation extent observed in TA stained with H&E (Fig. 5A,B). No changes were detected between experimental groups in GAS muscle, likely due to already high basal inflammation extent in this muscle in both strains (Fig. 6B,D). HO-1-dependent anti-necrotic action was detected also in diaphragm of mdx;HMOX1Pax7Ind both in untrained and exercised transgenic mice in comparison to mdx counterparts (Fig. 6B,E).
Fig. 6
HMOX1 induction in SCs attenuates skeletal muscle necrotic events upon physical exercise. A-E Necrosis assessment in tibialis anterior (TA), gastrocnemius (GAS) and diaphragmae (DIA) tissue sections of mdx and mdx;HMOX1Pax7Ind mice. (A) Representative scans of TA. Scale bar: 500 μm. B Representative images of necrotic events in TA, DIA and GAS. Scale bar: 100 μm. Semiquantitative analysis in (C) TA, (D) GAS and (E) DIA. Immunofluorescent staining of IgG and IgM (green) (n = 3–8). Data is presented as mean± SEM. # p < 0.05—Two-way ANOVA variation; * p < 0.05 by Two-way ANOVA with Tukey’s post hoc test
留言 (0)