
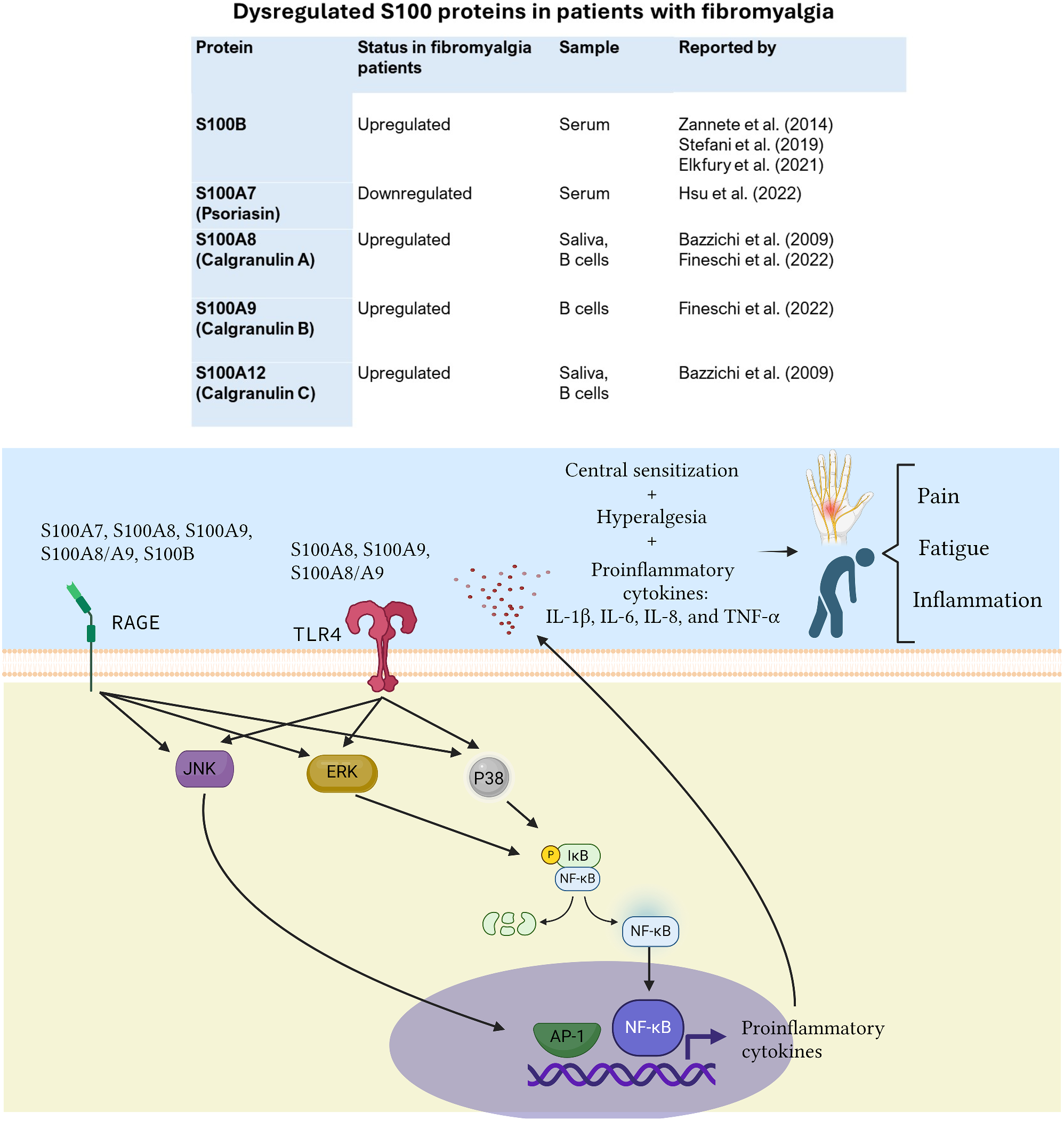
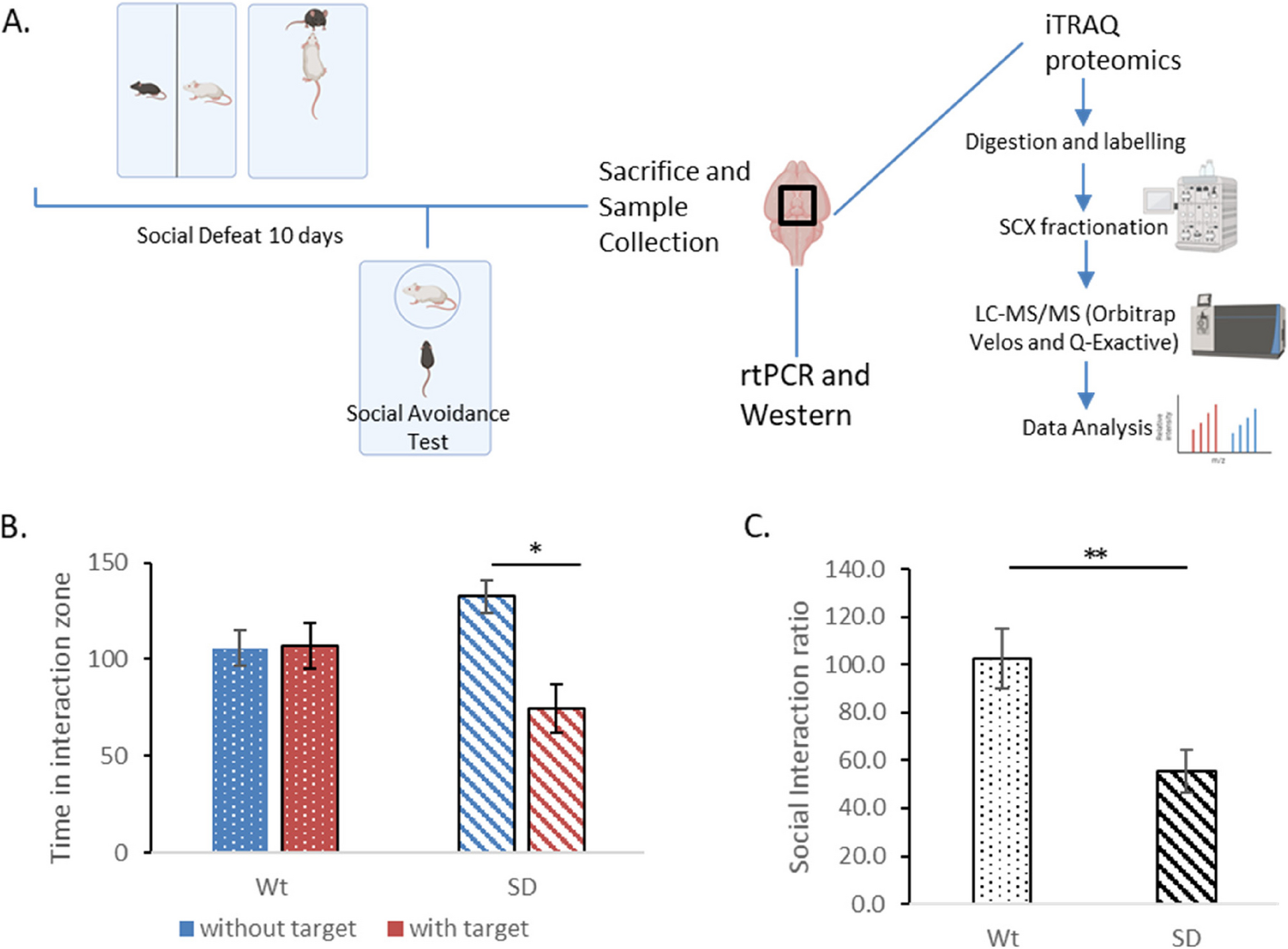
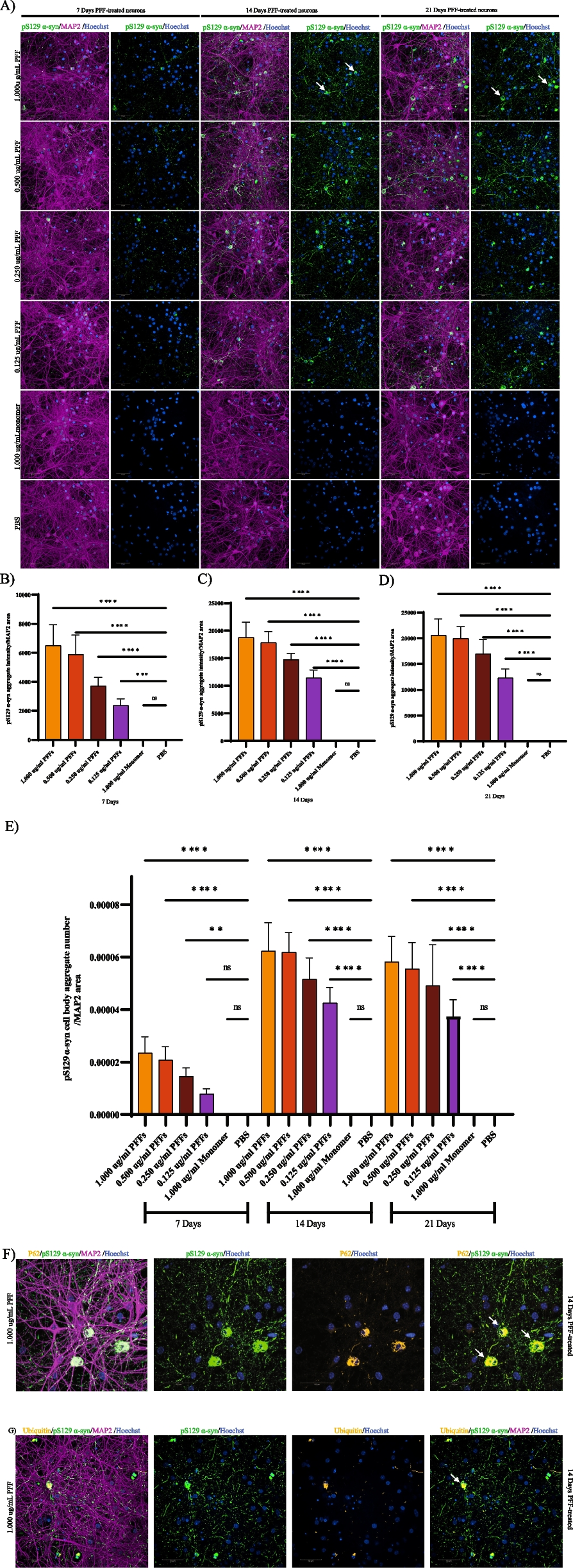
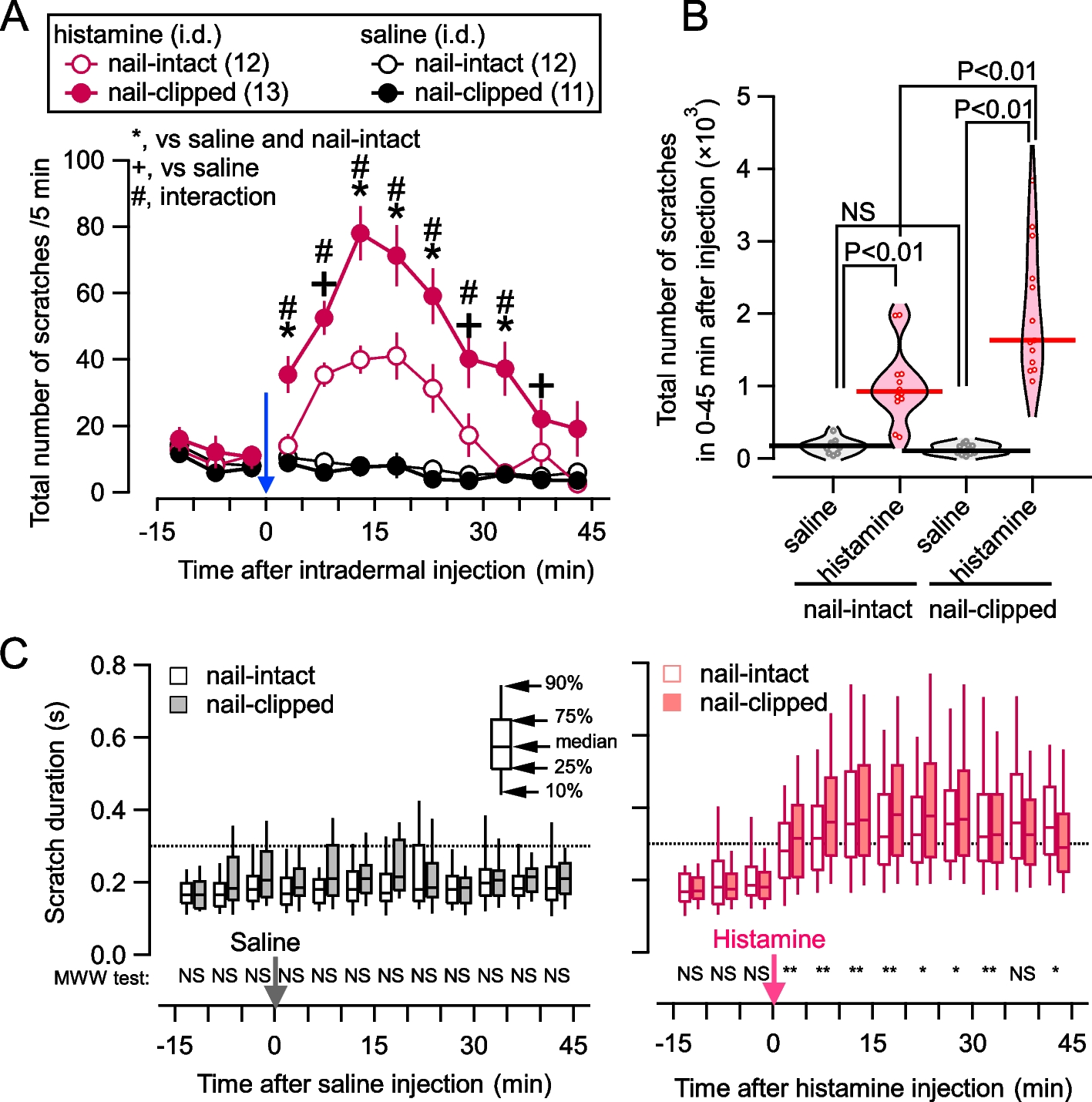
記住我

Chronic psychological stress has emerged as a key determinant in the development and progression of various neurological complications, including anxiety disorders, dementia, and depression [1,2,3]. Recent studies have shown that prolonged exposure to psychological stress can disrupt normal neurobiological processes, leading to structural and functional changes in key brain regions involved in emotional regulation, cognition, and memory [4, 5].
Autophagy is a highly regulated intracellular mechanism responsible for maintenance of cellular homeostasis by removing damaged organelles, protein aggregates, and other cellular debris [6]. In contrast to this well-established homeostatic and cytoprotective roles of autophagy, autophagy can also function as one of the modes of programmed cell death through the degradation of key cellular components [7]. Autophagic death has been implicated in various physiological and pathological contexts, including the response to cellular stressors such as nutrient deprivation, hypoxia, or exposure to toxins [8].
We previously reported that chronic restraint stress (CRS) leads to cognitive deficits and mood dysregulation by inducing autophagic death of adult hippocampal neural stem cells (NSCs), highlighting the potential impact of stress on the regenerative capacity of the brain [9].
In the past, programmed cell death has not been considered as the mechanisms underlying stress-induced hippocampal damage and cognitive impairments, due to the lack of the evidence of apoptosis [10, 11]. Consistent with those previous reports, CRS did not induce caspase 3 activation or chromosomal DNA fragmentation, well-known apoptotic markers [9]. Instead, CRS increased autophagy flux in adult hippocampal NSCs and NSC-specific, inducible deletion of Atg7, one of the key autophagy genes, prevented CRS-induced death of adult hippocampal NSCs and preserved cognitive functions [9]. These findings suggest that CRS suppresses adult hippocampal neurogenesis by triggering autophagic death of adult hippocampal NSCs and provide a new perspective on neurobiological mechanisms and potential therapeutic strategies for stress-related neurological disorders. However, it remains unknown whether other models of psychological stress elicit similar effects on adult hippocampal neurogenesis through autophagic death of NSCs.
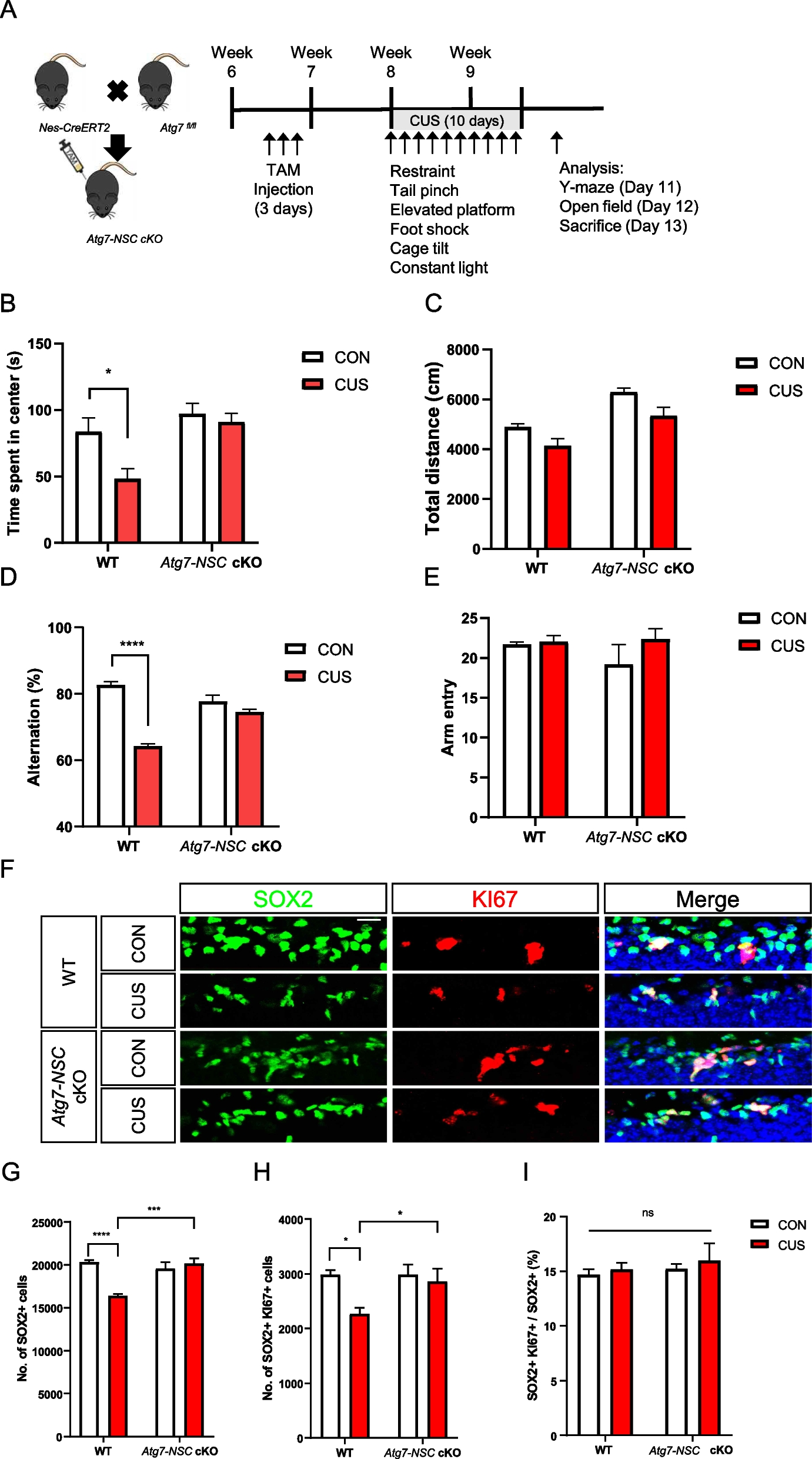
To address this question, mice were subject to chronic unpredictable stress (CUS) for 10 days (Table 1) and we examined the cellular and behavioral changes of mice caused by CUS and the role of Atg7 in CUS-induced loss of adult hippocampal NSCs. First, to generate mice with an inducible NSC-targeting Atg7 conditional knockout (Atg7-NSC cKO) mice, Atg7fl/fl mice were crossed with nestin-CreERT2 (Nes-Cre). In our previous study, we induced heterozygous ablation of Atg7 to avoid compounding effects of prolonged suppression of autophagy [9]. To induce Atg7 conditional knockout at the adult stage, 7-week-old Atg7fl/+ (WT) or nestin-CreERT2: Atg7fl/+ (Atg7-NSC cKO) mice were intraperitonially (i.p.) injected with tamoxifen (TAM) for 3 days and CUS was performed 1 week later. For 10 days, mice were exposed to one of six types of stresses daily, as detailed in Table 1. To investigate whether NSC-specific deletion of Atg7 keeps brain function intact, adult neurogenesis-dependent hippocampal functions were tested. After CUS, WT mice spent shorter time in the center of the open field in the open field test (Fig. 1B) and open arms in the elevated maze test (Figure S1A) and showed less spontaneous arm alternation in the Y-maze test (Fig. 1D) and decreased discrimination index in the novel object location test (Figure S1C), suggesting anxiety-like behaviors and spatial memory impairment by CUS. However, stressed Atg7-NSC cKO mice were resistant to CUS and did not show these neurobehavioral alterations (Fig. 1B, D, Figure S1A, C). There was no difference between WT and Atg7-NSC cKO groups in the total traveled distance in open field test (Fig. 1C) and total number of arm entries in Y-mase (Fig. 1E) and elevated plus maze tests (Figure S1B). To measure the death of NSCs, we counted the number of total (SOX2-positive) and proliferating NSCs (KI67/SOX2 double-positive) after CUS (Fig. 1F). CUS significantly decreased the numbers of total (Fig. 1G) and proliferating (Fig. 1H) NSCs in WT mice. Of note, the ratio of the proliferating NSCs was not affected by CUS, confirming that the decrease in the number of NSCs was due to death of NSCs rather than inhibition of proliferation (Fig. 1I). In contrast, Atg7 ablation blocked death of NSCs and prevented CUS-induced decrease in the number of NSCs (Fig. 1F-I). To demonstrate an increase in autophagy flux in vivo, we performed stereotaxic injection of Nestin promoter-driven mRFP-EGFP-MAP1LC3B lentivirus in mouse hippocampus, as we previously performed [9]. This tandem MAP1LC3B construct demonstrated a higher ratio of autolysosomes and an increase in the total number of MAP1LC3B puncta in WT hippocampal NSCs following CUS (Figure S1D-E), suggesting an increase in autophagy flux. However, these increases were not observed in Atg7-NSC cKO mice (Figure S1D-E). In contrast to the induction of autophagy flux, we were unable to detect activated caspase-3 (cleaved caspase-3, c.CASP3, Figure S1F) and TUNEL signals (Figure S1G) following CUS. As positive control, staurosporine (STS) was stereotaxically injected for the induction of CASP3 activation and DNA damage (Figure S1F-G). These results indicate the reduction of hippocampal NSCs by autophagy but not apoptosis, following CUS.
Table 1 A schedule for chronic unpredictable stress for 10 daysFig. 1
NSC-specific deletion of Atg7 prevents CUS-induced anxiety-like behavior, memory deficit, and the reduction in the number of adult hippocampal NSCs. A Experimental schedule of CUS given for 10 days. B, C Open field test. Time spent in the center (B) and total distance (C) in the open field test. D, E Y-maze test. Arm alternation (D) and total arm entry (E) in the Y-maze test. F Representative images of SOX2 and KI67 staining in the subgranular zone of the hippocampus. Scale bar: 20 μm. G-I Quantification of SOX2+ and KI67+ cells. Number of SOX2 + cells (G), SOX2 + KI67 + cells (H) and the ratio of SOX2 + KI67 + to SOX2 + cells (I) (n = 6 or 7 per group). *P < 0.05, *** P < 0.001, **** P < 0.0001. ns, not significant
Chronic stress can increase the risk of developing depression and relapse by inhibiting the production of new neurons [12]. Current treatments for depression and stress-related neurological disorders requires long-term medication and are not effective for all patients, and some patients may have drug resistance or experience side effects [13]. Therefore, it is imperative to find new treatment targets to improve the treatment effectiveness of depression and stress-related neurological disorders.
Our results demonstrate that Atg7-NSC cKO mice are functionally intact by prevention of autophagic death of adult hippocampal NSCs. Understanding the role of autophagic death in the context of chronic stress can provide insights into the mechanisms underlying stress-induced brain disorders and potentially guide the development of new therapeutic strategies to mitigate their impact.
留言 (0)