
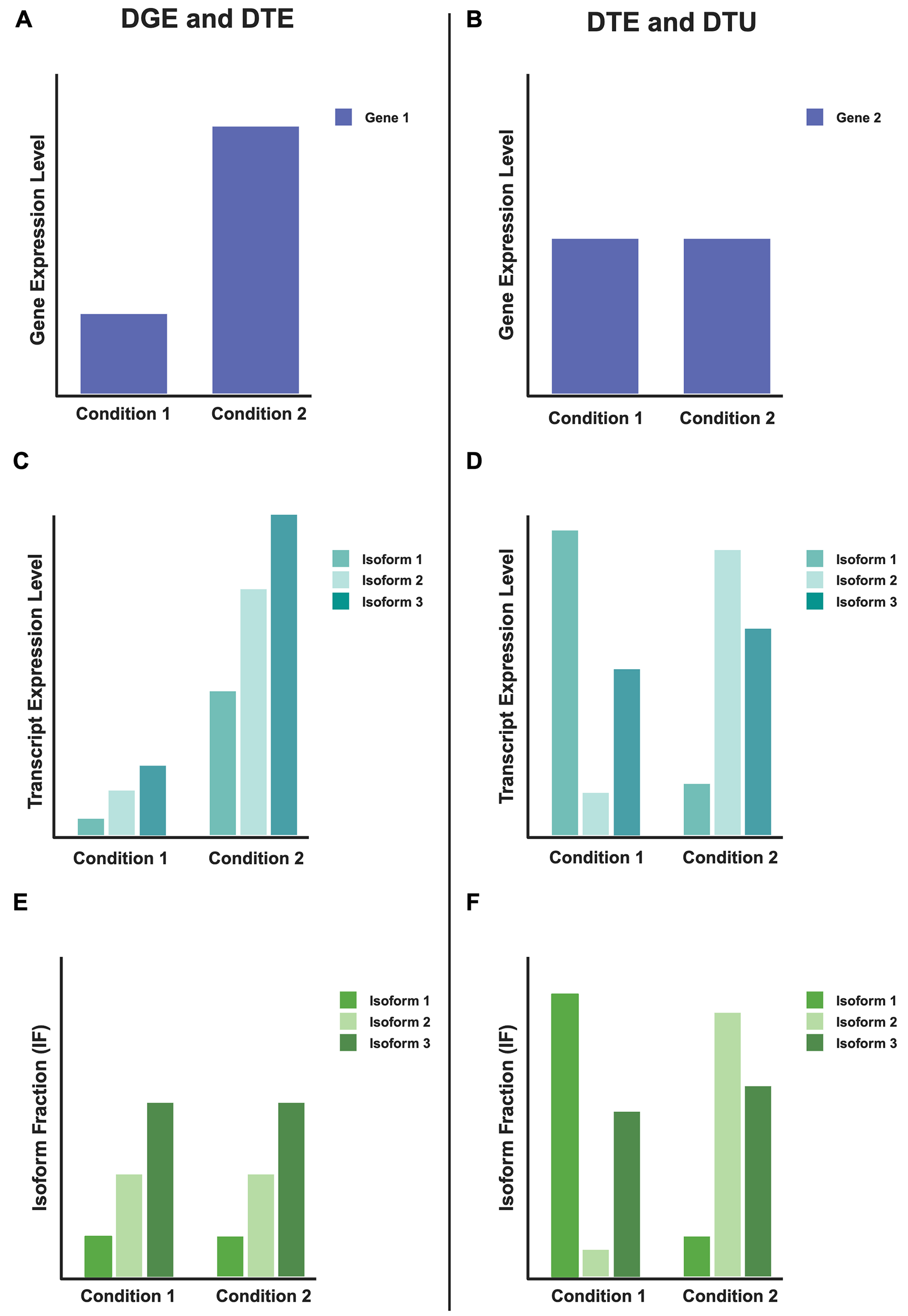
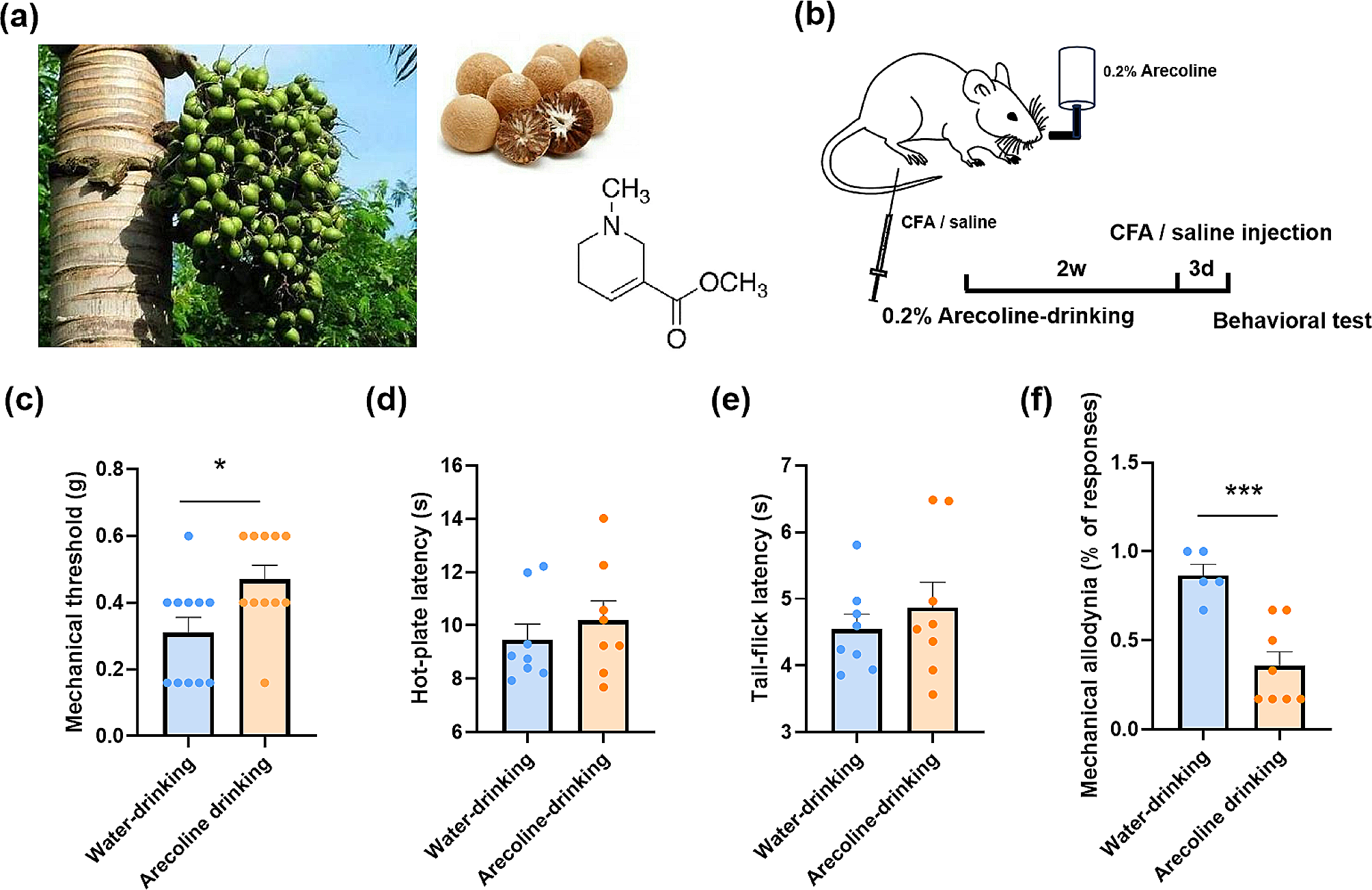
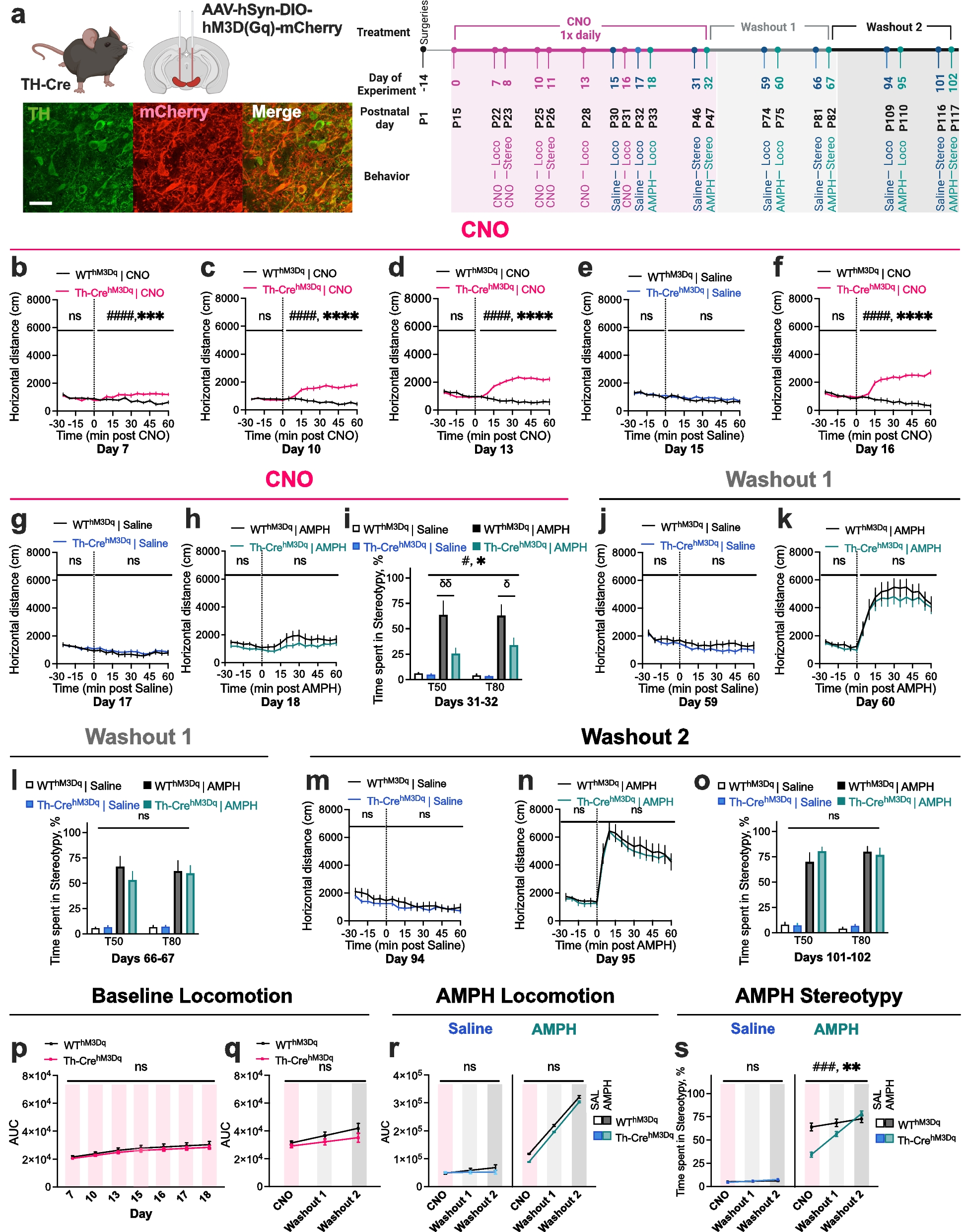
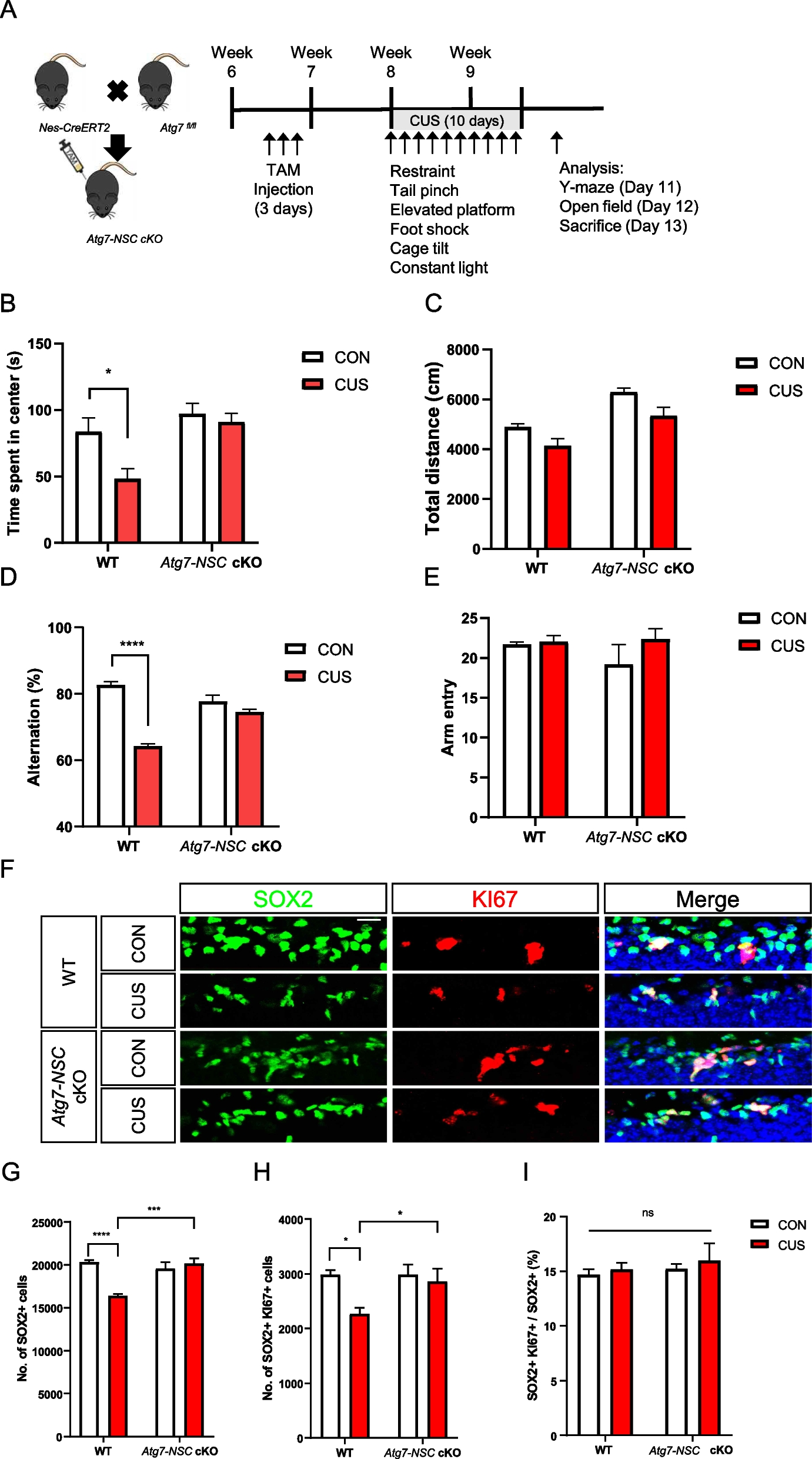
記住我

One of the main features of fibromyalgia is the heightened sensitivity to pain, known as hyperalgesia. This hyperalgesia could be explained in part by central sensitization, which refers to an abnormal amplification of pain signals within the central nervous system. S100 proteins, particularly S100A8 and S100A9, are significantly upregulated in inflammatory conditions and have been shown to mediate inflammatory responses and pain perception. Thus, S100 protein-mediated inflammation could be a key element in the pathophysiology of FM. This review presents the S100 proteins that have been reported as dysregulated in patients with fibromyalgia and discusses their potential contribution to the pathophysiology of FM.
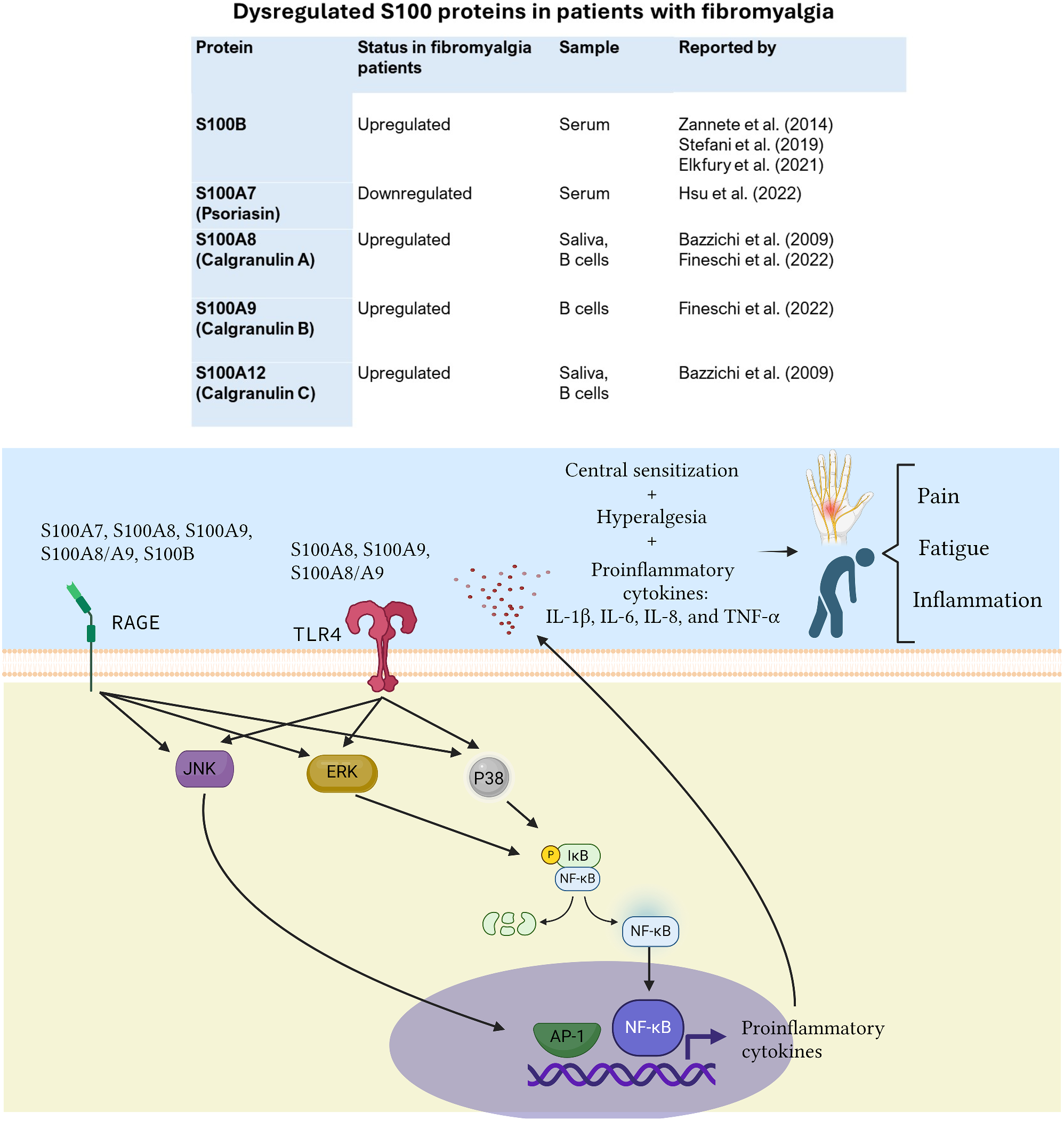
Dysregulated S100 proteins in patients with fibromyalgia: clinical reportsIn patients with fibromyalgia, S100A8 (calgranulin A) and S100A12 (calgranulin C) were found to be upregulated in saliva samples [1], while the expression of S100A8 and S100A9 was found to be increased in peripheral B cells of these patients [2]. S100B is the most studied S100 protein in patients with this chronic condition, and it has been found to be significantly elevated in serum of women with several chronic pain conditions, including fibromyalgia [3, 4]. In contrast, the serum levels of S100A7, were found to be lower in FM patients than in controls [5]. The disruption of these S100 proteins in individuals diagnosed with fibromyalgia suggests that these proteins play a pivotal role in the pathogenesis of this disease.
Potential mechanisms of S100 proteins in fibromyalgia pathophysiologyWhen released extracellularly, S100 proteins can act as damage-associated molecular patterns (DAMPs) and contribute significantly to inflammatory pain. It is well established that S100A7, S100A8, S100A9, S100A8/A9, S10012, and S100B bind to RAGE, while S100A8, S100A8, and S100A8/A9 dimer bind to toll-like receptor (TLR4). Both RAGE and TLR4 are expressed in cells such as macrophages and signal through JNK, ERK, and p38 to activate the transcription factors AP-1 (Activating Protein-1) and NF-κB (nuclear factor kappa B), which in turn induce the release of proinflammatory cytokines [6,7,8]. It has been demonstrated that increased proinflammatory cytokines, such as IL-1β, IL-6, IL-8, and TNF-α, are associated with the development of neuropathic pain and fatigue. IL-6 has been shown to alter the resting membrane potential threshold and cause hyperexcitability in neurons, which is secondary to phosphorylation of the sodium channel Nav1.7. This results in an increase in the transmission of pain signals. Furthermore, IL-6 has been implicated in the development of pain related to changes in neuronal plasticity [9, 10]. In a previous study, it was reported that serum concentrations of IL-6 and IL-8 were increased in FM patients. These cytokines exhibited a positive correlation with clinical scores on the Fibromyalgia Impact Questionnaire (FIQ), suggesting that IL-6 and IL-8 may have additive or synergistic effects in perpetuating the chronic pain experienced by FM patients [11]. Taken together, it is possible that the activity of S100 proteins is associated with the generation of pain through the release of proinflammatory cytokines (Fig. 1).
Fig. 1
The potential role of S100 proteins in the pathophysiology of fibromyalgia. S100A8, S100A9, S100A8/A9, S10012, and S100B bind to RAGE, while S100A8, S100A8, and S100A8/A9 dimer bind to toll-like receptor (TLR4). Both RAGE and TLR4 signal through JNK, ERK, and p38 to activate the transcription factors AP-1 (Activating Protein-1) and NF-κB (nuclear factor kappa B), which in turn induce the release of proinflammatory cytokines. These proinflammatory cytokines, coupled with the presence of hyperalgesia and central sensitization, may contribute to inflammation, fatigue and chronic pain
S100 proteins have also been considered as potential biomarkers for both CNS damage and brain tumors [11]. In addition, S100 proteins have been associated with changes in neuronal electrical properties, suggesting a possible role in neuronal sensitization [12]. Interestingly, S100A4 has also been implicated in the modulation of neuropathic pain in rodent models [13]. It is important to note that chronic pain is linked to neural plasticity, which refers to changes in pain transmission and perception in the CNS [14]. Therefore, the increased levels of S100 proteins, particularly S100B (together with BDNF), may be associated with changes in neuronal plasticity, which translates to chronic pain in patients. This is particularly relevant when discussing hyperalgesia and central sensitization. The increased perception of pain caused by hyperalgesia could be partially explained by central sensitization, which amplifies pain signals within the CNS. Central sensitization causes temporal, spatial and threshold alterations in pain sensitivity and is a clear example of the contribution of the CNS to the generation of pain hypersensitivity. Central sensitization enhances the function of neurons and circuits in nociceptive pathway, resulting in an increased response of the somatosensory nervous system in response to inflammation and neural injury [15]. When considering the contribution of S100 proteins to the inflammatory state, the pathophysiology of FM becomes more complex, since we must consider that these patients are potentially more susceptible to inflammatory processes. The pathophysiology of fibromyalgia involves a non-virtuous cycle between hyperalgesia, central sensitization, and S100 protein-mediated inflammation. This cycle significantly contributes to one of the most characteristic symptoms of fibromyalgia, which is chronic widespread pain. Figure 1 illustrates the potential role of S100 proteins in the pathophysiology of fibromyalgia.
留言 (0)