
記住我



Neuropathic pain (NP) represents a frequent concomitant condition in individuals suffering from spinal cord injury (SCI). Available treatments are ineffective and frequently associated with significant side effects. Identification of mechanisms underlying SCI-associated NP and subsequent development of targeted interventions represent important aims to ultimately help improve the quality of life of SCI survivors.
Over the years, several neuroanatomical regions have been identified as potential sites of NP initiation and transmission. There is the injury site characterized by an unpredictable degree of spinal cord damage that varies with respect to preserved ascending sensory pathways—the spinothalamic and the lemniscal tracts. Recently, secondary alterations in the peripheral nervous system (PNS) below injury level have come more into focus. We wondered whether hyperexcitable nociceptors are no longer properly controlled by appropriate mechanoreceptor input because of SCI-induced paraparesis or tetraparesis with resulting inability to stand or walk, thus depleting physiological sensory input from the periphery into the dorsal horn. Accordingly, a number of studies were conducted, which showed that sensorimotor activation through treadmill training in rodents with moderate thoracic contusion reduced pain behavior, associated with reversal of structural rearrangements below injury level in the dorsal horn, namely reduction of peptidergic nociceptor sprouting into laminae III to IV.34,41,44
If mechanoreceptors transmit sensory input provided by sensorimotor activation (eg, treadmill training) into the dorsal horn, they should keep by these means, based on the gate control theory of pain, nociceptor firing in check.1,13,30 Specific genetic ablation of tropomyosin receptor kinase B (TrkB) expressing low-threshold mechanoreceptors (LTMR) in mice undergoing SCI would allow us to address this hypothesis.
Electrophysiological characterization provides another window into the relevance of mechanoreceptors in NP induction/modulation. Hyperexcitability has previously been demonstrated in nociceptors after SCI.46 However, these findings were primarily based on dorsal root ganglia (DRG) cultures taken from SCI animals. While planning this study, such analyses have not been reported yet in ex vivo skin-nerve preparations from below injury level, considered to be a better approximation towards pathophysiological events in the living animal compared with cultures.
The relevance and comparability of animal SCI models in respect to the situation in human SCI subjects is very important. Of course, identical analyses are not applicable to human subjects for obvious reasons. However, quantitative sensory testing (QST), which stands for a battery of noninvasive psychophysical measures, allows assessment of the somatosensory system and—more specifically—hypersensitivity towards pain and mechanical stimuli,45 thus matching preclinical assessments.
Our main aim was to understand the contribution of components of the PNS, in particular mechanoreceptors, to induce and/or maintain SCI-mechanical allodynia after genetic ablation of mechanoreceptors and post-SCI in skin-nerve preparation electrophysiological examination. Findings were compared with noninvasive psychophysical measures (QST) reflecting the state of pain/touch hypersensitivity in SCI subjects up to 1 year after injury.
2. Materials and methods 2.1. Animal subjects and experimental groupsAdult female C57BL/6J mice (8-12 weeks old at experimental initiation; wild type; JANVIER LABS) of varying genotypes subdivided into different experiments weighing between 20 and 25 g were used in this project. Animals were housed in groups of 4 to 5 mice per cage on a 12/12-hour light/dark cycle with access to water and food ad libitum. To ensure optimal conditions for housing and behavioral testing, the facility was controlled for temperature (22°C ± 1°C) and humidity (45%-65%) on a daily basis. All experiments were planned and conducted according to the PREPARE and ARRIVE guidelines.36,42
For experimental purposes and to ensure that the same investigator can perform behavioral testing, animals were divided in different cohorts for each experiment. Data from the different cohorts were combined in the end for statistical analysis.
2.2. Spinal cord injuryAll contusion surgeries were conducted with the Infinite Horizon Impact Device (IH-0400 Impactor; Precision Systems & Instrumentation, Lexington, KY) adapted for the mouse spinal cord using a standard mouse steel-tipped impactor with a diameter of 1.3 mm.34,41 Before surgery, mice were deeply anesthetized with an intraperitoneal (i.p.) injection using a cocktail (2.5 mL/kg) containing ketamine (31.25 mg/kg), xylazine (1.58 mg/kg), and acepromazine (0.31 mg/kg). T9 laminectomy (corresponding to the T11 spinal level) was performed on mice of the injury group followed by a moderate contusion with a force of 50 kDyn. After contusion, the paravertebral muscle layers were sutured (4/0 terylene) and the skin incision was stapled. In sham mice, skin incisions and separation of paravertebral muscles to isolate the T9 vertebra was performed followed by suturing and stapling of the skin. Postsurgical care included analgesic medication, volume substitution, manual bladder voiding twice per day, and prophylactic antibiotic treatment to prevent bladder infection for 10 to 14 days. Because of the difficulty of emptying male bladders, infections, and post-SCI fighting, only female mice were used in the studies presented here.
All mouse experiments were conducted in accordance with the European Communities Council Directive (Directive 2010/63/EU amended by Regulation (EU) 2019/1010 and institutional guidelines) and approved by the local governing body (Regierungspräsidium Karlsruhe, Abteilung 3—Landwirtschaft, Ländlicher Raum, Veterinär-und Lebensmittelwesen', Germany (approval number: G-196/15)).
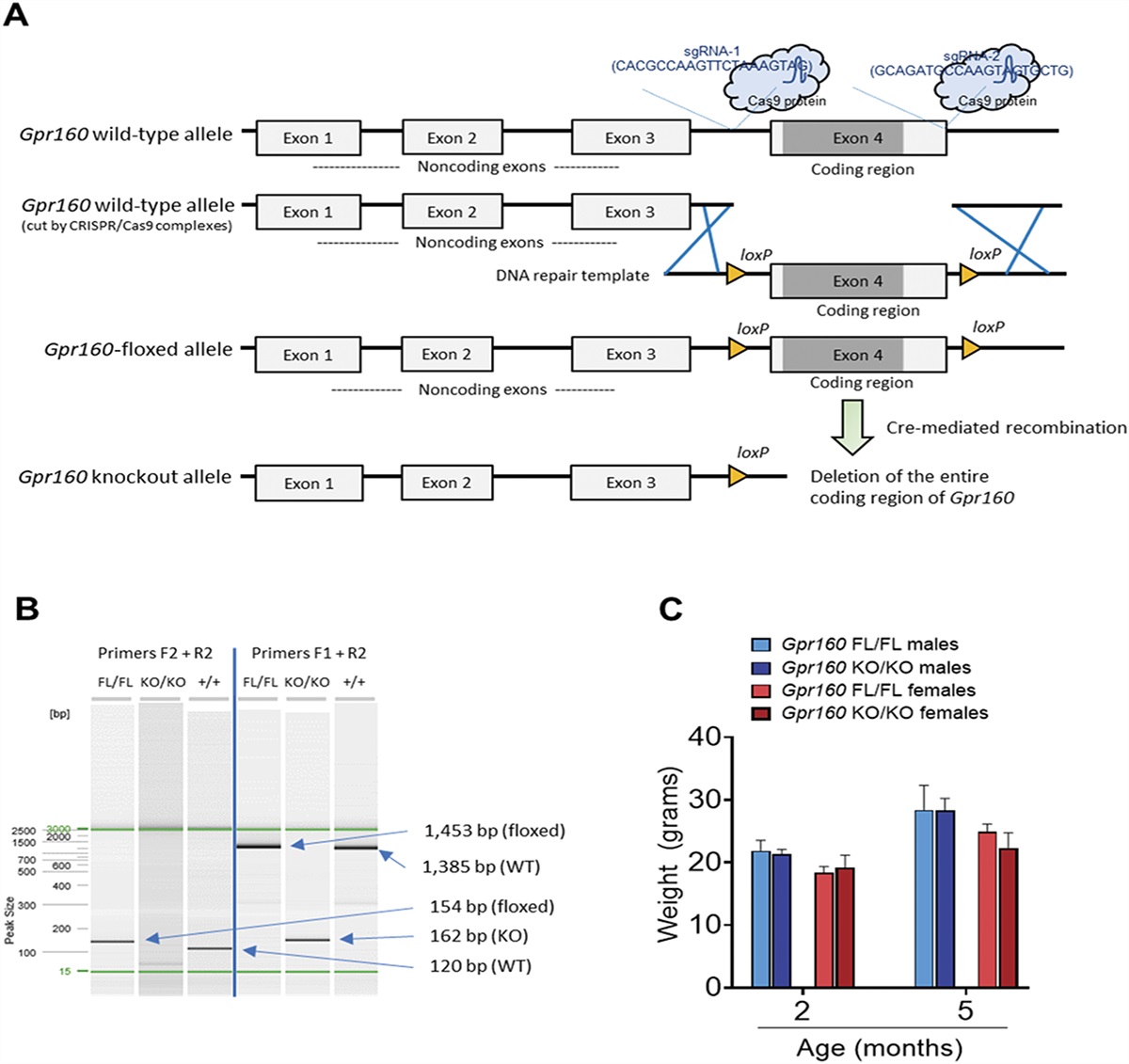
2.3. Transgenic animalsTo investigate the contribution of low-threshold mechanosensory neuronal subpopulations to SCI-induced below-level NP, heterozygous TrkBCreERT2::AviliDTR (TrkB inducible diphtheria toxin receptor (iDTR))12,22 in which mechanosensory neurons can be specifically ablated have been used. To generate heterozygous TrkBCreERT2::AviliDTR (TrkBiDTR) animals, C57BL/6-Tg(TrkB-CreERT2)1Phep/Embph (TrkBCreERT2) mice were crossed to C57BL/6-Tg(Avil-tm1-DTR)1Phep/Embph (AviliDTR) mice.
Administration of tamoxifen (75 mg/kg i.p. for 5 consecutive days) into TrkBiDTR mice specifically renders Avil-expressing TrkB+ sensory neurons susceptible to ablation by diphtheria toxin (DTX). Adult female TrkBiDTR (n = 19) mice were injected i.p. with 40 μg/kg DTX (Sigma, D0564) with a second dose after 72 hours. In TrkBiDTR animals, DTX was administered 14 days after the last tamoxifen injection unless described otherwise. Control animals were injected i.p. with 100 μL saline twice in the same time frame. TrkBiDTR animals were used to investigate the effect of ablation after SCI when mechanical allodynia has developed (SCI n = 17; +Tam/+DTX n = 9; +Tam/+saline n = 6) or on development of mechanical allodynia (before SCI n = 18; +Tam/+DTX n = 10; +Tam/+saline n = 8).
2.4. Behavioral testingMice were habituated to the individual testing set-up for 2 to 3 days for at least 1.5 hours per day before behavioral testing and for 30 to 60 minutes in each set-up on testing days. All behavioral testing was performed in awake, unrestrained mice by the same investigator blinded to group identity.
2.4.1. Basso mouse scaleRecovery of motor function was assessed using the Basso Mouse Scale (BMS) for locomotion as described before.34,41 The motor function was scored 1 day post-injury to confirm an adequate injury and weekly starting 7 days post-injury. Each hind paw was scored individually and averaged to get a single score for each animal per testing day. A score of 0 reflects complete hind limb paralysis without ankle movement, whereas a score of 9 indicates normal locomotion. The BMS was used to evaluate sensorimotor function to ensure proper recovery of motor function for sensory testing, which requires the animals to show weight support (BMS ≥ 3).
2.4.2. Mechanical sensitivity testingTo evaluate changes in mechanical sensitivity after injury and ablation of specific sensory neuronal subpopulations, von Frey hair filaments with ascending diameter and force (0.04, 0.07, 0.16, 0.4, 0.6, and 1.4 g; Touch Test Sensory Evaluators; North Coast Medical, Gilroy, CA) were applied to the hind paws of mice as described before.34,41
Starting with the smallest filament (0.04 g), the plantar surface of each hind paw was stimulated as previously described, resulting in 10 stimuli per filament per mouse.34,41 To avoid stimulus-induced sensitization of the hind paw, up to 12 animals were tested in parallel and hind paws were stimulated in an alternating manner. Mechanical sensitivity towards each filament is defined as paw withdrawal frequency (number of positive responses divided by the number of stimuli) and expressed as the average for both hind paws. Each animal was tested on 2 consecutive days before surgery and the mean was defined as the preoperative baseline response rate.
To investigate nocifensive response duration (removal, licking, and shaking) of a subcohort of wild-type mice (SCI n = 4, sham n = 4), video recording of the response towards the 0.16- and 1.4-g von Frey filament was performed preoperatively and 7 days post-injury (iPhone XR, 1080p HD, 240 fps, slow-motion function, 4.16 milliseconds/frame). Response duration was defined as the time until the hind paw was placed down again.
2.4.3. Place escape/avoidance paradigmThe place escape/avoidance paradigm (PEAP) is based on the assumption that animals escape and/or avoid an aversive mechanical stimulus. Animals have the active choice between a naturally preferred dark environment or pain relief in a more aversive light environment.3 Here, it was used to address the aversive quality of pain and thereby supraspinal processing of nociception as previously described.41 In short, animals were placed in the middle of the chamber, and the time spent on the dark side of the chamber was measured. An initial 10-minute exploration phase (baseline) without any stimulus was followed by a 15-minute testing phase (3× 5-minute blocks) in which the hind paws were stimulated using a 0.16-g von Frey filament every 15 seconds when the mouse was present on the dark side of the chamber. The left and right hind paws were stimulated in an alternating manner.
2.4.4. Thermal sensitivity testingThermal sensitivity was assessed according to Hargreaves' method using the plantar test (Hargreaves Apparatus; Ugo Basile) as previously described.34,41 In short, an infrared laser beam (190 ± 1 mW/cm2) was presented to the plantar surface of the hind paws and the time (sec) until withdrawal was recorded, resulting in the response latency to the heat stimulus.24 To avoid tissue damage, a 15-second cut-off time, as well as a delay of at least 1 minute between 2 trials for the same paw and mice, was introduced. In each animal, the mean of 4 trials for the left and right hind paws was used to express thermal sensitivity, and testing was always performed after mechanical sensitivity was assessed. Similar to von Frey testing, the mean latency of 2 consecutive testing days before surgery or any other intervention determined the baseline of the animals (preoperatively; preintervention).
2.5. Tissue processing and morphological analysisAll animals were euthanized with an overdose of anesthesia cocktail (ketamine, xylazine, and acepromazine) and transcardially perfused with 0.9% saline followed by 4% paraformaldehyde (PFA) in 0.1 M phosphate buffer.34 The spinal column was dissected, postfixed in 4% PFA for 1 hour at room temperature, and briefly rinsed with ddH2O followed by a 5-minute washing in 0.1 M phosphate buffer before the tissue was cryoprotected in 30% sucrose in 0.1 M phosphate buffer at 4°C. The lesion site (T11; if applicable), the L4 to L6 spinal cord, as well as L4 to L6 DRG were identified using anatomical landmarks,25 embedded in Tissue-Tek O.C.T compound (Sakura Finetek, Staufen, Germany) followed by serial sectioning (spinal cord, 16 and 25 µm coronal; DRG, 10 µm) directly on to slides as previously described.34,41
2.5.1. Eriochrome Cyanine stainingThe lesion size of injured animals was analyzed by quantifying the spared tissue at the lesion epicenter. Two consecutive series were stained for myelin using Eriochrome Cyanine (EC) as previously described.34,41 Tissue sparing at the lesion epicenter (smallest area of spared white matter) was quantified by outlining areas with light and dark blue staining and expressed as the percent of cross-sectional area calculated by dividing the white matter area by the total cross-sectional area. Imaging and analysis was performed blinded to group identity using ImageJ.
2.5.2. ImmunohistochemistryImmunohistochemical labeling was performed on slides with serial sections of the lumbar spinal cord (L4-L6) and the corresponding L4 to L6 DRG. Slides were dried for 30 minutes at room temperature, encircled with a liquid blocker, and rinsed in 0.1 M Tris-buffered saline (TBS). Sections were blocked and permeabilized by incubating in TBS/0.25% Triton TX-100/5% donkey serum followed by incubation with primary antibodies diluted in TBS/0.25% Triton TX-100/1% donkey serum overnight at 4°C. On day 2, sections were incubated with secondary antibodies and 4′,6-diamidino-2-phenylindole (DAPI), rinsed, dried, and cover-slipped with Fluoromount G (Southern Biotechnology Associates, Birmingham, AL).41 For each animal, images of the left and right dorsal horn of 3 labeled sections with an intersection distance ranging from 700 to 1400 µm were taken at ×20 magnification with the same resolution, lens aperture, and exposure time using a XC30 camera mounted on an Olympus BX53 microscope with epifluorescence illumination and appropriate filter cubes. Lumbar DRG (L4-L6) were imaged bilaterally from each animal at ×10 magnification using the above-described set-up, and images from 3 sections per DRG (140 µm apart) were analyzed.
The following primary antibodies were used: rabbit anti-calcitonin gene-related peptide (CGRP) (Immunostar, Hudson, WI, 24112; 1:200), rabbit anti-protein kinase Cγ (PKCγ [C19]; Santa Cruz Biotechnology, Dallas, TX, SC-211; 1:200), guinea pig anti-NeuN (Millipore, Darmstadt, Germany, ABN90; 1:2000) and goat anti-human HB-EGF (R&D Systems, Minneapolis, MN, AF-259-NA; 1:40). Secondary antibodies were Alexa Fluor 488 and Alexa Fluor 594 donkey anti-rabbit and donkey anti-goat (all Life Technologies, Carlsbad, CA; 1:300), Alexa Fluor 594–conjugated streptavidin (Jackson ImmunoResearch Ely, United Kingdom; 1:300), and Alexa Fluor 488 donkey anti-guinea pig (Jackson ImmunoResearch; 1:300).
2.5.3. Quantification of labeling density in the spinal cordThe termination pattern of peptidergic fiber and nonpeptidergic fiber was analyzed in lamina III to IV of the dorsal horn as previously described.41 In short, an analysis box was placed in the center of the dorsal horn adjoining the ventral border of lamina Iii, which was identified by PKCγ labeling. The dorsoventral extent of lamina IIi was used as a reference for the dimensions of the region of interest (ROI) analysis box. Images were processed by setting a labeling threshold minimizing background and accurately reflecting CGRP- and Tomato-positive fiber labeling. Labeling density was expressed as percentage of positive labeling within the analysis box, and the values for 3 sections per animal were averaged. Quantification of labeling density was performed blinded to group identity using ImageJ.
2.5.4. Quantification of ablated DRG neuronsIn TrkBiDTR L4 to L6 DRG quantification of HB EGF+ in NeuN+ neurons was used to evaluate the proportion of neurons expressing diphtheria toxin receptor (DTR) (DTR+/NeuN+).
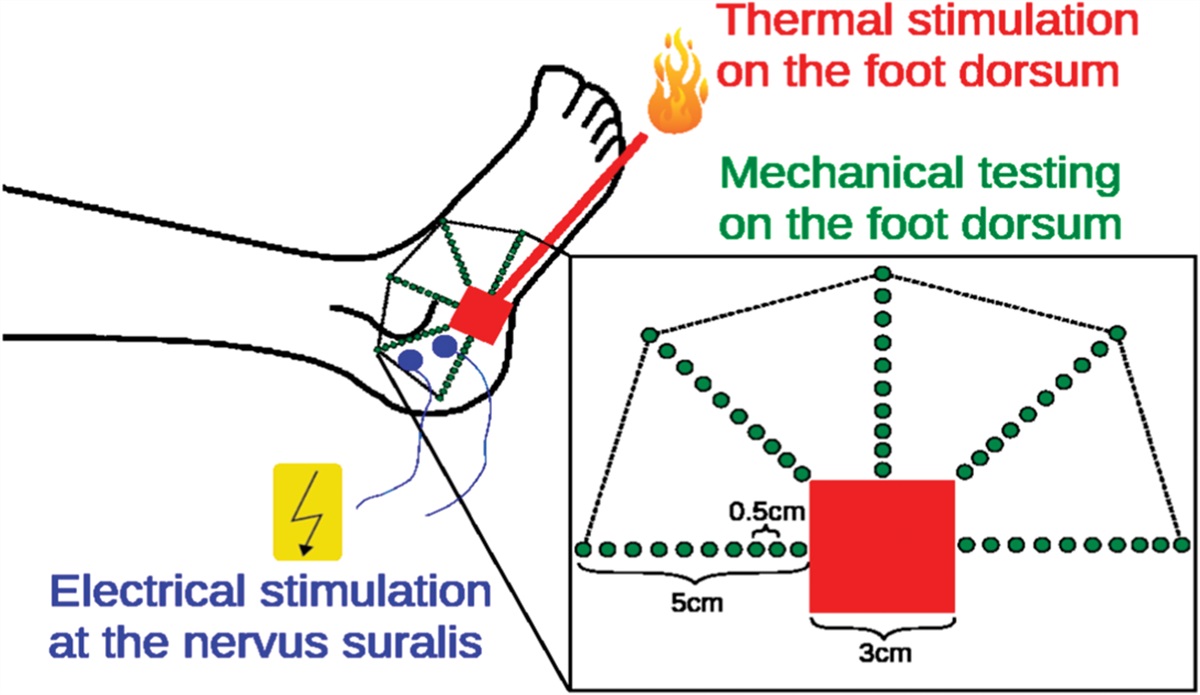
2.6. Ex vivo skin nerve recordingsSkin-nerve recordings were performed on 10- to 12-week-old SCI or Sham mice killed with CO2 followed by cervical dislocation. The hind paw skin was dissected free together with the sural nerve and was placed in an organ bath chamber that was perfused with 32°C-warm synthetic interstitial fluid (SIF) consisting of 108 mM NaCl, 3.5 mM KCl, 0.7 mM MgSO4, 26 mM NaHCO3, 1.7 mM NaH2PO4, 1.5 mM CaCl2, 9.5 mM sodium gluconate, 5.5 mM glucose, and 7.5 mM sucrose at a pH of 7.4. The sural nerve was led through a small hole into the adjacent mineral oil filled recording chamber. The nerve was teased into thin bundles that were laid on a silver wire recording electrode connected to a differential amplifier (Digitimer, modules NL104, NL125/NL126) and nerve fiber activity was recorded with the Powerlab 2 4SP system and Labchart 7.1 software (AD Instruments, Sydney, Australia). The receptive fields of single nerve fibers were located by manually probing the skin using von Frey hair filaments. To distinguish between Aβ fibers, Aδ fibers, and C fibers, action potentials were evoked by electrical stimulation of the receptive fields, and the axonal conduction velocities were calculated by dividing the distance between the stimulation electrode and the recording electrode by the delay between the onset of electrical stimulation and the arrival of the action potential at the recording electrode. Nerve fibers with a conduction velocity >10 m/second were classified as Aβ fibers, fibers with a CV between 1 and 10 m/second as Aδ fibers, and fibers with a CV <1 m/second as C fibers. To further distinguish between nociceptors and mechanoreceptors, the mechanical activation thresholds were determined with von Frey hair filaments. Finally, to determine the action potential firing rates and adaptation patterns at different stimulus strengths, the receptive fields were stimulated with ramp-and-hold stimuli applied with a linear piezo actuator (Nanomotor; Kleindiek, Reutlingen, Germany) equipped with a force measurement system (FMS-LS; Kleindiek, Reutlingen, Germany) to measure the exact force of the applied stimulus.
2.7. Human spinal cord injury studyIncluded participants (n = 17) were identified and recruited from July 2016 to June 2021 by convenient sampling. All of them provided written informed consent. The human SCI study was conducted within the framework of the European Multicenter Study about Spinal Cord Injury (EMSCI) at the SCI center, Heidelberg University Hospital (ClinicalTrials.gov register-no. NCT01571531; https://emsci.org). The study protocol was approved by the local ethical review board (S-188/2003). Inclusion criteria for the EMSCI study are acute traumatic or ischemic SCI. Furthermore, the initial assessments must be performed within the first 6 weeks after injury to be eligible for inclusion. Exclusion criteria were a nontraumatic cause of SCI other than spinal cord ischemia, impaired capabilities in respect to cooperation or giving informed consent, medical history of polyneuropathy, and concomitant traumatic brain injury.9
Sensory function was assessed in SCI subjects by both the International Standards for Neurological Classification of SCI (ISNCSCI) and highly standardized QST.27,38 Quantitative sensory testing represents a comprehensive protocol for assessing the integrity of the sensory system along the complete nervous system. For the classification of existing spontaneous pain problems in individuals with SCI, all participants received the clinical assessment EMSCI Pain Assessment Form (EPAF) that is in accordance with international guidelines and recommendations for the clinically documentation and assessment of pain in SCI.48 Within EMSCI, all study participants underwent recurrent comprehensive clinical examinations in predefined time windows (up to day 40, day 70-98, day 150-186, and day 300-546) within the first year after injury.9 For this study, only data of 3 timepoints (1, 3, and 12 months) were used. Six individuals did not complete the final timepoint.
In consideration of the methodological approach for the animal model, special focus was set on the evaluation of the mechanical detection threshold (MDT) representing a test for Aβ fibers (mechanoreceptors) and the mechanical pain threshold (MPT) to function as indicator for Aδ fibers (nociceptors).4,21,51 Although MDT was performed via modified von Frey hair filaments (contact area 0.5 mm in diameter; Optihair2-Set, Marstock Nervetest, Germany) applying forces from 0.25 to 512 mN, starting with 8 mN, MPT was assessed by means of (weighted) pinprick examination (contact area 0.25 mm in diameter) applying forces from 8 to 512 mN, starting with 8 mN.4
Dedicated areas being tested as to alterations of sensory function were the dermatomes L4 (medial right shin, midway between knee and ankle) and L5 (dorsum of the foot, third metatarsal phalangeal joint). The rationale for choosing L4/L5 was based on related literature reporting a frequent localization of NP in these regions.10,17,20,49
Evaluation and analysis of QST data was done based on standard reference MDT and MPT values provided by 180 healthy subjects (respecting test site, sex, and age, split by decades) as provided by the German Research Network on Neuropathic Pain (DFNS) by means of a QST reference database.32,33
2.8. Statistical analysisBehavioral results were analyzed by repeated-measures (RM) 2-way analysis of variance (ANOVA) to reveal overall group and timepoint differences, without assumption of sphericity (Geisser–Greenhouse corrections). Significant group differences were followed by Sidak pairwise comparisons. Significant timepoint differences between baseline vs post-SCI or baseline vs post-DTX were followed by post hoc Fisher least significant difference (FLSD) test.
For experiments in which transgenic animals have been used, the lesion size, fiber density in the lumbar spinal cord, and changes in DRG neurons (ablated neurons) were only compared between saline injected control SCI and ablated SCI group and consequently analyzed by unpaired t test.
Electrophysiological data were analyzed with multiple χ2 tests and multiple Mann–Whitney tests.
All data are presented as mean ± SD, except for skin-nerve preparation data, which are presented as mean ± SEM. Statistical analysis was done using Prism 9 software (GraphPad Software Inc, La Jolla, CA) with an alpha level of 0.05 for significance.
Quantitative sensory testing data were processed and analyzed using the software “eQUISTA” (Casquar GmbH, Bochum, Germany). Accordingly, results are expressed as Z-transformed standard values (“Z-scores”). Values greater than 1.96 and lower than −1.96, indicating ∼2 SDs from healthy matched controls findings (with 95% confidence interval), meaning hypersensitivity or hyposensitivity to the respective stimulus.38,39
3. Results 3.1. Pre-injury ablation of tropomyosin receptor kinase B–expressing mechanosensory neurons reduces neuropathic pain behavior after mouse contusion spinal cord injuryWe hypothesized that low-threshold mechanoreceptors (LTMRs), responsible for light touch, mediate beneficial sensory input provided by natural locomotor activity or in an enhanced setting such as treadmill training.16,28,29,44 Therefore, we investigated the contribution of TrkB-expressing LTMRs (identified as Aδ-LTMRs, D-hair, and rapidly adapting Aβ mechanoreceptors, Aβ-RAMs12) to SCI-induced neuropathic pain behavior represented by mechanical allodynia by ablating TrkB-expressing LTMRs in TrkBCreERT2:AviliDTR (TrkBDTR) mice.
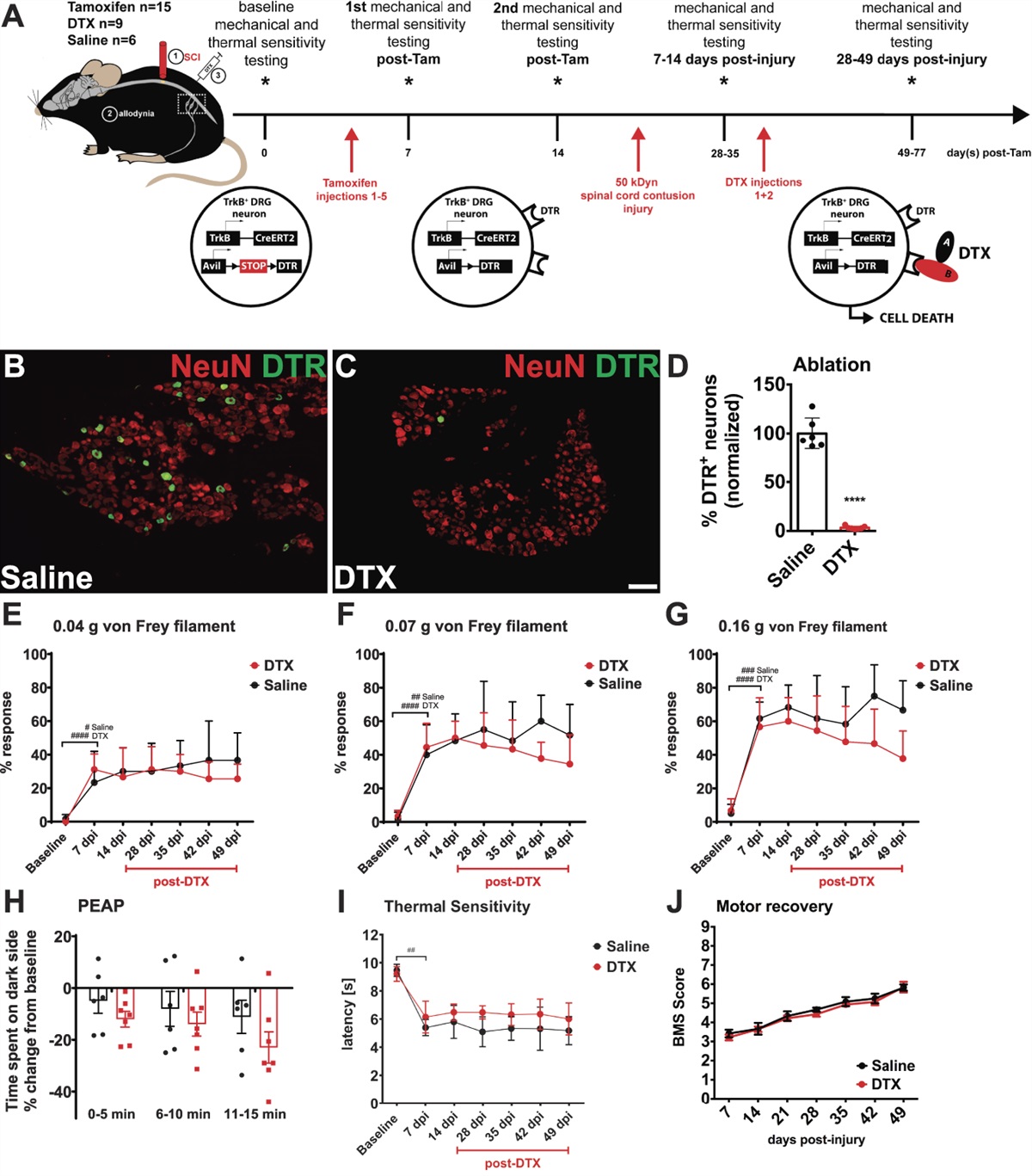
First, to determine the contribution of TrkB-expressing sensory neurons to established signs of SCI-induced neuropathic pain, respective neurons were genetically ablated 14 days after thoracic contusion SCI using diphtheria toxin (DTX), when mechanical allodynia and thermal hyperalgesia have already developed (Fig. 1A). Before SCI, tamoxifen administration did not affect mechanical and thermal sensitivity (data not shown), and as expected, all SCI animals developed mechanical allodynia and thermal hyperalgesia at 7 days post-injury (Figs. 1E–G,I, 2-way RM ANOVA, time differences P < 0.0001). Genetic ablation resulted in a 97% reduction in TrkB+/DTR+ neurons (Figs. 1B–D; t test P < 0.0001 DTX vs saline). Despite the high ablation efficiency, no significant effect of post-SCI ablation on mechanical hypersensitivity was observed. Statistical analysis indicated no overall group differences comparing the response rates for the small-diameter von Frey hair filaments (0.04 g-0.16 g) between saline-injected control and ablated SCI animals (Figs. 1E–G, 2-way RM ANOVA group: 0.04 g, P = 0.54; 0.07 g, P = 0.27; and 0.16, g P = 0.06). In line with this observation, supraspinal processing assessed by the PEAP also showed no difference between groups in response to the 0.16-g filament (Fig. 1H, 2-way RM ANOVA P = 0.23 for group differences).
 Figure 1.:
Figure 1.: Post-SCI ablation of TrkB-expressing mechanoreceptors. (A) Schematic of experimental design. (B) Representative lumbar (L4-L6) DRG neurons of saline-treated control animals stained for HB-EGF shows robust expression of DTR in a subset of sensory neurons. (C) In ablated TrkBDTR animals, DTR expression is almost completely absent (scale bar, 100 μm). (D) Quantification (t test, P < 0.0001; DTX, n = 7; saline n = 6, L4-L6 bilaterally). (E–G) SCI-induced mechanical hypersensitivity to small-diameter light touch eliciting von Frey hair filaments (0.04, 0.07, and 0.16 g, 2-way RM ANOVA, time differences P < 0.0001, baseline vs 7 dpi FLSD #P < 0.05, ##P < 0.01, ###P < 0.001, ####P < 0.0001), but DTX-mediated ablation of TrkB+ sensory neurons (28-49 dpi) did not change SCI-induced mechanical hypersensitivity. (H) The Place Escape Avoidance Paradigm (PEAP, 0.16 g) performed at the end of the study shows that both groups spent more time on the light side of the chamber (2-way RM ANOVA P = 0.23 for group differences). (I) Injured TrkBDTR animals of both groups develop thermal hypersensitivity assessed by the Hargreaves test (2-way RM ANOVA, baseline vs 7 dpi P < 0.0001, FLSD ##P < 0.01, group differences P = 0.02, no significant Sidak pairwise comparisons). (J) Except for the SCI-induced motor deficits scored by the Basso Mouse Scale (BMS), no further motor deficits are observed after post-SCI ablation of TrkB+ sensory neurons. Mean ± SD for all graphs. 2-way RM ANOVA, 2-way eepeated-measures analysis of variance; DTR, diphtheria toxin receptor; DTX, diphtheria toxin; FLSD, Fisher least significant difference; FLSD, Fisher least significant difference; HB-EGF, heparin binding EGF-like growth factor ; SCI, spinal cord injury; TrkB, tropomyosin receptor kinase B.
Post-SCI ablation did not affect response rates measured using larger diameter innocuous von Frey hair filaments (Suppl. Fig. 1A, available at https://links.lww.com/PAIN/B970, 0.4 g: 2-way RM ANOVA P = 0.17) but did have significant group differences for noxious stimuli (0.6 g and 1.4 g) with inconsistent or nonsignificant pairwise comparisons at each timepoint (Suppl. Fig. 1B-C; 2-way RM ANOVA, group differences 0.6 g, P = 0.04; 1.4 g, P = 0.01). TrkB-expressing LTMR ablation did not affect motor recovery (Fig. 1J). After SCI, significant thermal hypersensitivity (Hargreaves method) was observed for all SCI animals irrespective of ablation (Fig. 1I, 2-way RM ANOVA, baseline vs 7 dpi P < 0.0001, group differences P = 0.02, no significant Sidak pairwise comparisons). No significant differences in white matter sparing (lesion size) were observed between saline-injected controls and ablated SCI mice (Suppl. Fig. 2A, available at https://links.lww.com/PAIN/B970; t test P = 0.52). Taken together, these results suggest that TrkB-expressing sensory neurons do not play a role in already developed and stably present mechanical allodynia after SCI.
Second, to determine the role of TrkB-expressing sensory neurons in the initiation of SCI-induced neuropathic pain, neurons were ablated before SCI. This resulted in an effective reduction of >90% of DTR+ neurons in lumbar DRG (L4-L6) (Figs. 2B–D; t test P < 0.0001 DTX vs saline control group). Without injury, no differences in mechanical or thermal sensitivity were observed (Figs. 2, Suppl. Fig. 1D-F, available at https://links.lww.com/PAIN/B970). All SCI animals of the saline-injected control group showed mechanical hypersensitivity at 7 days post-injury compared with pre-SCI values using small-diameter von Frey filaments (Figs. 2E–G). Interestingly, a significant group effect was observed (Figs. 2E–G; 2-way RM ANOVA group differences: 0.04 g, P = 0.004; 0.07 g, P = 0.008; 0.16 g, P = 0.013). However, TrkB-ablated SCI animals (DTX group) showed a lower response rate than the control group only at 21 days post-injury, indicating a small but significant effect of pre-injury ablation of TrkB-expressing sensory neurons on SCI-induced mechanical allodynia. In line with the lack of group differences to the 0.16-g von Frey hair filament at the end of the experiment, the PEAP indicated SCI-induced mechanical allodynia in both groups (Fig. 2H, 2-way RM ANOVA group differences P = 0.49). Pre-SCI ablation did not affect response rates measured using larger diameter (0.4-1.4 g) von Frey hair filaments (Suppl. Fig. 1D-F, 2-way RM ANOVA, group differences 0.4 g, P = 0.11; 0.6 g, P = 0.11; 1.4 g, P = 0.13). Both, thermal sensitivity (Fig. 2I, P = 0.14) and motor recovery (Fig. 2J) were not influenced. Additionally, differences in sensory behavior of saline-injected and ablated SCI animals cannot be attributed to difference in white matter sparing (lesion size) (Suppl. Fig. 2B, available at https://links.lww.com/PAIN/B970, t test P = 0.69). Taken together, only pre-injury—not post-injury—ablation of TrkB-expressing mechanosensory neurons reduces SCI-induced mechanical allodynia.
 Figure 2.:
Figure 2.: Pre-SCI ablation of TrkB-expressing mechanoreceptors. (A) Schematic of experimental design. (B) Saline-injected control animals show robust DTR expression in lumbar (L4-L6) DRG neurons. Compared with control animals, DTR+ neurons are almost completely ablated in DTX-injected animals (C) (scale bar, 100 μm). (D) Quantification (t test P< 0.0001; DTX, n = 9; saline n = 7, L4-L6 bilaterally). (E–G) SCI-induced mechanical hypersensitivity to small-diameter filaments (0.04, 0.07, and 0.16 g, 2-way RM ANOVA, time differences P < 0.0001, post-DTX vs 7 dpi FLSD #P < 0.05, ##P < 0.01, no significant (ns) difference between baseline and post-DTX) is significantly altered in ablated animals, but inconsistently (2-way RM ANOVA group differences: 0.04 g, P = 0.004; 0.07 g, P = 0.008; 0.16 g, P = 0013, Sidak pairwise comparisons *P < 0.05). The response rate towards the 0.16-g von Frey hair filament (G) of ablated animals is not significantly different from control animals anymore at the end of the experiment 28 dpi. (H) This is confirmed by supraspinal processing of nociception using the PEAP (0.16 g) where both groups equally spent more time on the light side of the box. (I) DTX treatment does not affect pre-injury or post-injury thermal sensitivity, but SCI did induce thermal sensitivity in both groups (2-way RM ANOVA, time differences P < 0.0001, post-DTX vs 7 dpi FLSD ##P < 0.01, ####P < 0.0001). (J) SCI induces significant motor deficits in all animals scored by the Basso Mouse Scale (BMS), and ablation of TrkB+ sensory neurons does not induce further motor deficits. Mean ± SD for all graphs. DTX, diphtheria toxin; FLSD, Fisher least significant difference; PEAP, place escape/avoidance paradigm; SCI, spinal cord injury; TrkB, tropomyosin receptor kinase B.
Regardless of the ablation timing (post- or pre-SCI), calcitonin gene-related peptide (CGRP) density in laminae III to IV of the lumbar dorsal horn was not reduced in respective transgenic SCI mice compared with the non-ablated SCI control group (Figs. 3A–F). Therefore, SCI-induced maladaptive structural changes regarding sprouting of CGRP-expressing fibers were not influenced by ablation of TrkB-expressing sensory neurons and existed while mechanical allodynia was present.
 Figure 3.:
Figure 3.: Peptidergic nociceptive fiber changes in the dorsal horn after TrkB-expressing mechanoreceptor ablation. Lumbar (L4-L6) spinal cord sections of injured saline control (A and D) and DTX-treated injured (B and E) TrkBDTR mice stained for CGRP (scale bar, 100 μm). Representative images indicate the SCI-induced increase in CGRP-labeling density in deeper laminae (III-IV) of the spinal dorsal horn in all groups. (C and F) Quantification of the CGRP-labeling density in laminae III-IV of the lumbar (L4-L6) dorsal horn shows no difference comparing the injured saline-injected control group with injured DTX-injected animals but a significant increase compared to previous sham animals (dotted line) (mean ± SD) (post-SCI; saline n = 6; DTX n = 7) (pre-SCI; saline n = 7; DTX n = 9). CGRP, Calcitonin gene-related peptide; DTX, diphtheria toxin; SCI, spinal cord injury; TrkB, tropomyosin receptor kinase B.
3.2. Experimental contusion spinal cord injury induces nociceptor, but not mechanoreceptor, hyperexcitabilityTo identify the key neuronal population (nociceptors or mechanoreceptors) driving mechanical allodynia as a correlate of NP behavior, ex vivo skin-nerve preparations were investigated.
Single-unit teased fiber recordings from ex vivo hind paw skin-nerve preparations revealed that C- and Aδ-fiber nociceptive sensory afferents displayed decreased mechanical activation thresholds 7 days post-injury (Figs. 4A–E). Thus, a significantly higher proportion of C-fiber nociceptors responded to forces smaller than 125 mN (Fig. 4E, multiple χ2 tests). Moreover, C-fiber nociceptors fired significantly more action potentials in response to mechanical ramp-and-hold stimuli (Fig. 4F, multiple Mann–Whitney tests) and exhibited continuous action potential firing after removal of the stimulus (Figs. 4G–I, multiple Mann–Whitney tests). These afterdischarges were also observed in Aδ-fiber nociceptors (Figs. 4G and H). Additionally, a significantly higher proportion of Aδ-fiber nociceptors responded to forces at 17.5 and 22.5 mN (Fig. 4C, multiple χ2 tests); however, they did not fire more action potentials in response to mechanical ramp-and-hold stimuli (Fig. 4D, multiple Mann–Whitney tests). Most importantly, no significant changes in the mechanical activation thresholds or the action potential firing rates were observed after SCI in the 3 major classes of low-threshold mechanoreceptors (rapidly adapting (RA) Ab-LTMRs, slowly adapting (SA) Ab-LTMRs, and D hairs (DH) Ad-LTMRs, Figs. 4J–M). Therefore, a midthoracic contusion SCI induces hyperexcitability of peripheral nociceptive fibers but not mechanoreceptors of the hind paw.
 Figure 4.:
Figure 4.: Mechanoreceptor and nociceptor activity after SCI. Single-unit teased fiber recordings from hind paw glabrous skin-nerve preparations show that A-fiber (A) and C-fiber nociceptors (B) have lowered activation thresholds 7 dpi T11 (50 kDyn) SCI in mice. This is quantitatively shown in (C) for A-fiber and (E) for C-fiber nociceptors responding to forces smaller than 125 mN (multiple χ2 tests). While following SCI A-fibers (D) do not fire more frequently, C-fibers (F) significantly fire more frequently in response to mechanical ramp-and-hold stimuli. Example traces exhibit continuous firing after removal of stimulus (150 mN denoted in gray area) in the SCI group (G). This is quantitatively shown for A-fiber (H) and C-fiber (I) nociceptors (multiple Mann–Whitney tests). The firing of RA-Aβ-LTMRs (J), SA-Aβ-LTMRs (K), and DH-Aδ-LTMRs (L) was not significantly altered by SCI. Additionally, LTMR activation thresholds were not altered by SCI (M). Mean ± SEM. * P < 0.05, ** P < 0.01. DH, D hairs; LTMRs, low-threshold mechanoreceptors; RA, rapidly adapting; SA, slowly adapting; SCI, spinal cord injury.
Does prolonged nocifensive behavior such as removal, licking, and shaking correlate with the extension of the firing past stimulus removal? Indeed, almost a doubling of the response duration of the SCI mice to the 0.16-g filament was observed at 7 days post-injury (Fig. 5A, t test P = 0.03). Also the response rate for these mice was significantly increased 7 days post-injury (Fig. 5B, 2-way RM ANOVA, group differences, P = 0.03). Therefore, the continued firing observed in nociceptors appears to be behaviorally expressed by prolonged nocifensive behavior in SCI mice.
 Figure 5.:
Figure 5.: Prolonged nocifensive behavior after experimental SCI. (A) T11 SCI mice showed prolonged nocifensive behavior (licking, shaking, and holding) compared with Sham mice when responding to 0.16-g von Frey hair filaments 7 dpi (slow-motion video analysis, unpaired 2-tailed t test, *P < 0.05). (B) The response rate of SCI rate was significantly higher than Sham mice at 7 dpi with 0.16-g von Frey hair filament (Sham n = 4, SCI n = 4, 2-way RM ANOVA, group difference P = 0.03, Sidak pairwise comparison ***P < 0.001). 2-way RM ANOVA, 2-way repeated-measures analysis of variance SCI, spinal cord injury.
3.3. Nociceptor hypersensitivity is observed in spinal cord injury subjects up to 1 year post-injuryThe best approximation towards the relevance in nociceptors vs mechanoreceptors in initiating/maintaining hyperexcitability and NP presentation, QST was used. Quantitative sensory testing was performed 1, 3, and 12 months post-injury in 17 conveniently sampled individuals with SCI varying in age (18-75 years), sex (male, n = 13; female, n = 4), cause of injury (traumatic, n = 11; ischemic, n = 5; or hemorrhagic, n = 1), initial classification according to the ASIA impairment scale (AIS D, n = 9; C, n = 3; B, n = 2; A, n = 3), and initial neurological level of injury (C2-T10, cervical = 9, thoracic = 8) (6 individuals did not complete the full study) (Suppl. Table 1, available at https://links.lww.com/PAIN/B970; Fig. 6). Not all SCI subjects could be tested at each timepoint and at all intended spinal segments (at 1 month, n = 14; at 3 months, n = 11; at 12 months, n = 8). The majority of sensory incomplete SCI subjects, in comparison with the healthy subject QST reference data bank, present with MPT Z-scores beyond the 1.96 SD margin (1 month: n = 10; 3 months: n = 9; 12 months: n =8; Fig. 6). Whereas, MDT Z-scores are mostly clustered within normal variation of 1.96 and −1.96, with some SCI subjects displaying Z-scores beyond the Z-score < −1.96 SD margin pointing towards reduced mechanosensation (5 being classified initially as exhibiting hypoesthesia at 1 month and 2 at 3 months and at 12 months in at least 1 dermatome). At 12 months, of the 8 remaining sensory incomplete individuals (AIS-B, C and D), 6 reported at-level (n = 2) or below-level (n = 4) NP.
 Figure 6.:
Figure 6.: Quantitative sensory testing in SCI subjects. Fourteen sensory incomplete SCI individuals (and 1 initial complete SCI subject) were examined 1, 3, and 12 months post-SCI for mechanical detection threshold (MDT), reflecting Aβ mechanoreceptors, and mechanical pain threshold (MPT), reflecting Aδ nociceptors. SCI individuals were measured at the L4 (right shin) and L5 (dorsum of right foot) dermatomes and found to be consistently altered compared with healthy database matched controls in MPT but not MDT (represented by Z-scores). Red dashed lines visually mark the Z-scores at 1.96 and −1.96, the cutoff for hypersensitivity or hyposensitivity, respectively. In the 12-month timepoint, open symbols represent at-level NP, lower filled symbols represent below-level NP, and fully filled symbols represent no NP. Mean ± SD. NP, neuropathic pain; SCI, spinal cord injury.
Comparing the MDT analysis with light touch (LT) assessment based on International Standards for Neurological Classification of SCI (ISNCSCI) (Fig. 6B; Suppl. Table 2, available at https://links.lww.com/PAIN/B970) yields concordant results. The majority of SCI subjects rate light touch neither normal nor absent equivalent to a score of 1 (Suppl. Fig. 3, available at https://links.lww.com/PAIN/B970). Accordingly in the MDT, several of the SCI subjects display reduced light touch sensation (tested with up and down method von Frey hair filaments). Pinprick assessment according to ISNCSCI vs MPT threshold results requires a more thorough explanation. The majority of SCI subjects displayed a pinprick score of 0 (ISNCSCI), indicating the subject either feels nothing or is not able to discriminate between sharp and dull stimuli. However, in the MPT (QST), the majority are depicted as hypersensitive towards pinprick stimuli (MPT Z-score >1.96).
In summary, sensory incomplete SCI subjects are characterized by early and persistent hypersensitivity towards nociceptor-mediated stimuli, whereas such changes could not be observed in respect to mechanoreceptor-mediated stimuli.
4. DiscussionResults from the present study improve our understanding of initiation and maintenance of NP signals in traumatic SCI. Neuropathic pain ameliorating effects of mechanoreceptor ablation depend on the timing of ablation in relation to the time of injury. Only pre-SCI mechanoreceptor ablation reduces pain behavior, suggesting a potentially NP-inducing role. Replicating previously published evidence, nociceptors become hyperexcitable early after SCI.
The finding that pre-SCI ablation of mechanoreceptors—not post-SCI ablation—reduces pain behavior, points toward an NP-inducing rather than an NP-maintaining effect. This is supported by the absence of mechanoreceptor hyperexcitability in hind paw skin-nerve preparation 7 days after thoracic contus
留言 (0)