
記住我



Neuropathic pain is a type of chronic pain due to lesions or diseases of the peripheral and/or central somatosensory nervous system.26,66 It is a worldwide problem that can affect productivity, lead to psychological comorbidities and financial burden, and overall negatively affect an individual's quality of life.21 These pain conditions are exceedingly difficult to treat; drugs currently used to manage pain in patients, such as opioids, have numerous side effects and high abuse liability.56 Therefore, there is a high priority to identify novel non–opioid-based analgesics to treat neuropathic pain. G-protein-coupled receptors (GPCRs) are the most desirable drug targets to treat various diseases because they regulate many processes, including pain, and roughly 40% of FDA-approved drugs target GPCRs.12,18
We recently reported that activation of the G-protein-coupled receptor 160 (GPR160) in the spinal cord by its ligand cocaine- and amphetamine-regulated transcript peptide (CARTp) contributes to the development of central sensitization associated with neuropathic pain states.75 Cocaine- and amphetamine-regulated transcript peptide is a neuropeptide expressed in areas involved in pain processing, including peptidergic C fibers, primary afferents that terminate in the superficial laminae of the spinal cord.42,43 Mechanistically, CARTp/GPR160-induced central sensitization is driven by activation of the mitogen-activated protein kinases (MAPKs) and extracellular signal-regulated kinase (ERK) and cAMP response element-binding protein (CREB).75 It is well-documented that MAPK activation in the spinal cord is an integral component of neuroinflammatory process and a mechanistic hallmark of various persistent pain states.15,38 In addition to our findings revealing increased expression of Gpr160 in 2 different animal models of nerve injury–induced neuropathic pain,75 there are also published transcriptomic analyses showing increased expression of Gpr160 in other models of pain. For example, genome-wide expression analysis found that Gpr160 is significantly upregulated in the spinal cord in a mouse model of spinal nerve ligation.4 Furthermore, Gpr160 is upregulated in the spinal cord in both male and female mice and rats that underwent spared nerve injury.31 In addition to animal models of neuropathic pain, Gpr160 is also expressed in patients with neuropathic pain. Indeed, a recent study identified Gpr160 as one of the top pain-associated genes upregulated in the dorsal root ganglion in a cohort of female patients with neuropathic pain who underwent thoracic vertebrectomy surgery for spinal reconstruction or tumor resection.58
Understanding the emerging contribution of GPR160 in pain processing has relied on the use of genetic and immunopharmacological approaches.75 The lack of small-molecule ligands for GPR160 hampers our understanding of the roles of CARTp/GPR160 signaling in pathophysiological settings. To begin addressing this void, we generated the first global Gpr160 KO mice using CRISPR-Cas9 (clustered regularly interspaced palindromic repeats-CRISPR-associated protein 9) genome editing technology and used these mice to further validate the role of CARTp/GPR160 in nociceptive behaviors.
In this study, we provide further evidence that CARTp causes behavioral hypersensitivities through GPR160 in a Gi/o-GPCR manner and supports GPR160 as a potential therapeutic target to treat neuropathic pain with limited cognitive and mood side effects.
2. Materials and methods 2.1. Experimental animalsMale and female Institute of Cancer Research (ICR) mice (7-8-week-old) were purchased from Envigo-Harlan Laboratories (Indianapolis, IN). ICR mice were used for experiments that did not involve Gpr160 KO mice. These include CARTp given intraplantar (i.pl.) (Figs. 3A and B) and pertussis toxin experiments (Figs. 2C and D, Figs. 3C and D). In experiments that involved the Gpr160 KO mice, the control animals used were the Gpr160-floxed mice that came from a C57BL/6 background. Mice were housed no more than 4 per cage with 12-hour light and 12-hour dark cycles in a controlled environment (regulated temperature, 20-22°C and 55%-65% humidity) with food and water available ad libitum. Animals were randomly assigned to the treatment group before the start of the experiment, and behavioral testing was performed between 9:00 am and 5:00 pm. Experimenters conducting the behavior tests and the injections were blinded to the treatment conditions. Experiments were performed in males and female mice, and similar behavioral data were obtained. Therefore, when males and females were used, the data were combined and the n values for each sex is indicated in the figure legends. All experiments were performed in accordance with the guidelines of the International Association for the Study of Pain (Seattle, MD) and the National Institutes of Health (Bethesda, MD) on laboratory animal welfare and approved by the Saint Louis University Animal Care and Use Committee (IACUC).
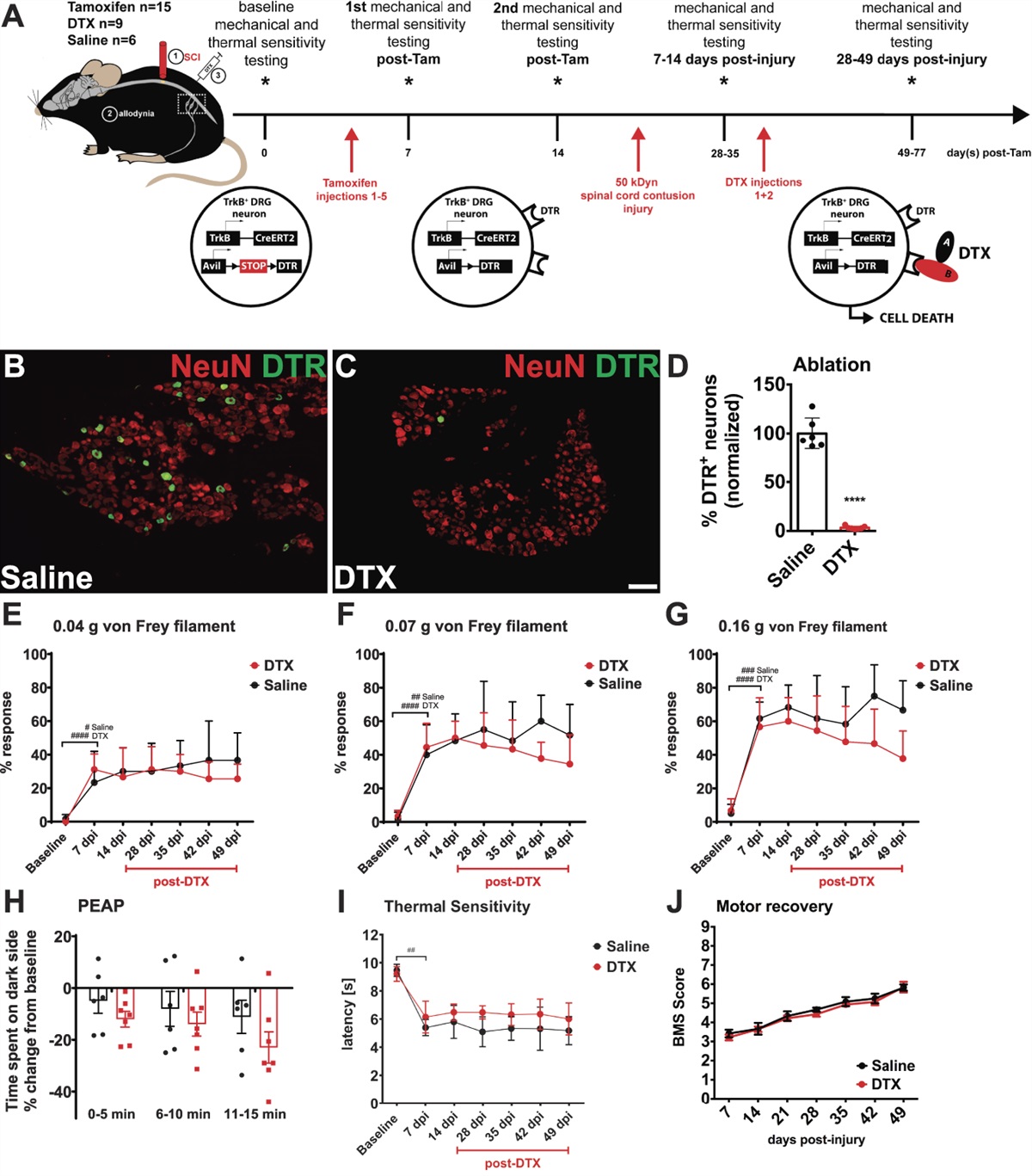
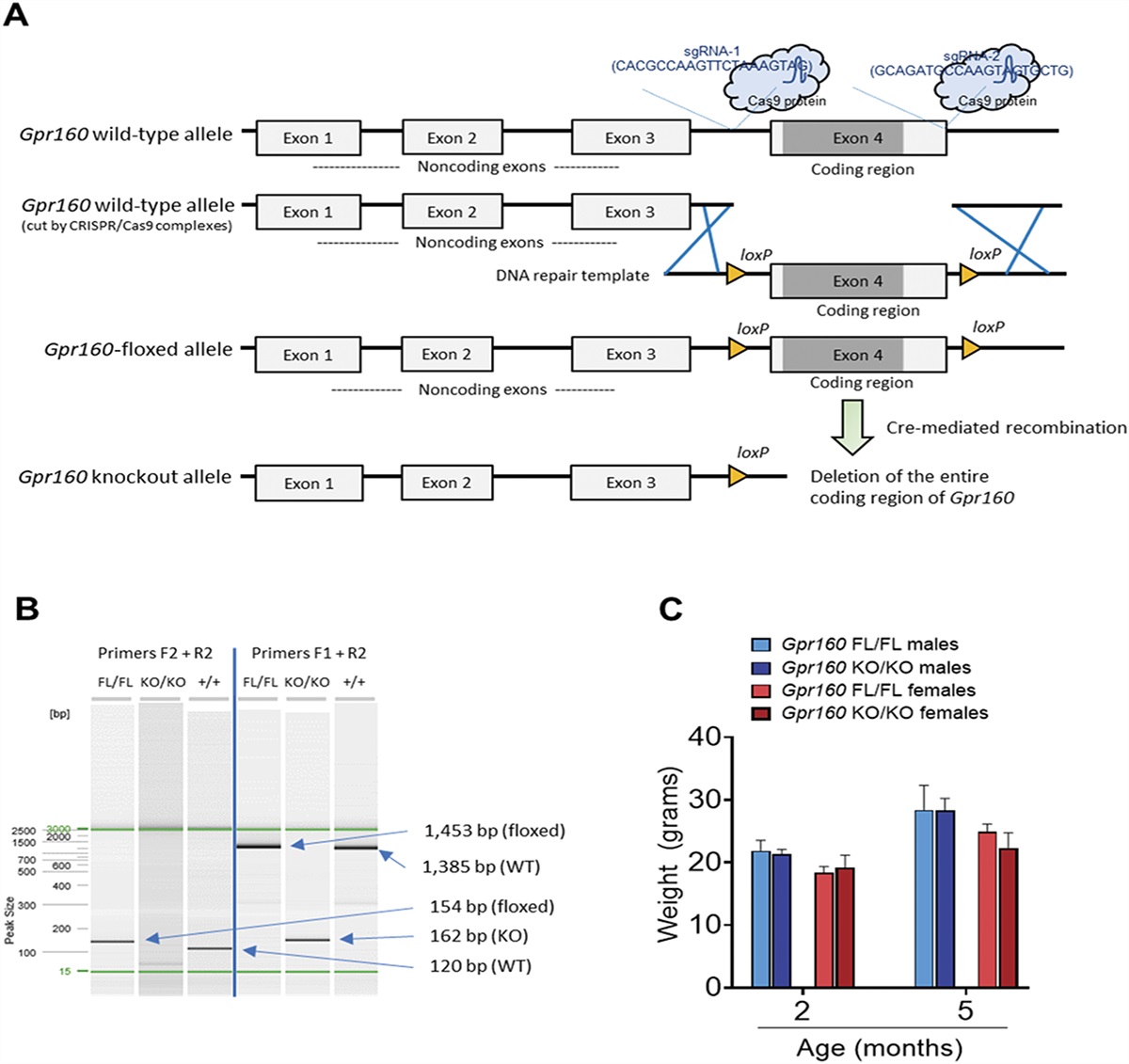
Gpr160-floxed animals were generated using CRISPR-Cas9 genome editing by the University of Missouri's Animal Modeling Core (Fig. 1A). All experimental procedures were approved by the University of Missouri's Institutional Animal Care and Use Committee and were performed according to the guidelines set forth in the Guide for the Use and Care of Laboratory Animals. C57BL/6 embryo donor females (3 weeks of age) and stud males (10 weeks of age) were purchased from Jackson Laboratory. CD-1 (ICR) surrogate females (8 weeks of age) were purchased from Charles River. Mice were housed in ventilated cages (Thoren) and kept on a 12:12 light–dark cycle. Food and water were available ad libitum. C57BL/6 mouse genomic sequence for G-protein-coupled receptor 160 (Gpr160) was obtained from Ensembl.org (assembly GRCm39). The CRISPR RGEN Tools Web site maintained by the Center for Genome Engineering Institute (Korea) was used to calculate off-target scores and design single-guide RNAs (sgRNAs) flanking the entire coding region of Gpr160. The CCTop Web site maintained by the Centre for Organismal Studies (Heidelberg) was used to calculate CRISPRater efficiency prediction scores.44 After designing, the sgRNAs were ordered as a chemically modified synthetic sgRNAs through Synthego (Supplemental Table 1, available at https://links.lww.com/PAIN/B974). The chemical modifications were 2′-O-methyl analogues and 3′ phosphorothioate internucleotide linkages at the first three 5′ and 3′ terminal RNA residues. A repair template was designed to introduce the desired loxP sites flanking the coding region of Gpr160 (Fig. 1A). The repair template was chemically synthesized by IDT as a long single-stranded DNA (lssDNA) (Supplemental Table 1, available at https://links.lww.com/PAIN/B974). The lssDNA was complementary to the target strand and contained asymmetrical homology arms (286 bp and 188 bp, respectively). A microinjection mix containing a final concentration of 50 ng/μL sgRNA (each), 50 ng/μL enhanced specificity Cas9 protein (Sigma Aldrich, St. Louis, MO), and 10 ng/μL lssDNA was made immediately before microinjections. In brief, CRISPR sgRNA-Cas9 ribonucleoprotein (RNP) complexes were formed by gently mixing the sgRNAs and Cas9 protein together and incubating at room temperature for 10 minutes. After RNP formation, the lssDNA repair template was added to the mixture. Pronuclear-injected zygotes were surgically transferred to pseudopregnant surrogate females the same day as microinjections. Offspring were delivered naturally and genotyped at 3 weeks of age. After DNA sequence confirmation of the correctly targeted allele, maintained animals were genotyped for the presence of each engineered loxP sites using polymerase chain reaction (PCR) primers flanking the individual sites. The 5′ loxP site was confirmed using PCR primers mGpr160-F1 (5′-GTCAACTAGGAATGCTTCCCTCCAC-3′) and mGpr160-R1 (5′-CAAGGCAGTCTGACGAGTACACAG-3′). A 171 bp amplicon is detected in wild-type samples, whereas a 205 bp amplicon is detected in floxed samples. The 3′ loxP site was confirmed using PCR primers mGpr160-F2 (5′-GGACAGGAACTGAAGCTCGAGTC-3′) and mGpr160-R2 (5′-TCCACAATGTTGTCGAGCAGGC-3′). A 120 bp amplicon is detected in wild-type (WT) samples, whereas a 154 bp amplicon is detected in floxed samples (Fig. 1B and Supplemental Figure 1, available at https://links.lww.com/PAIN/B974). Gpr160-floxed animals were then crossed with a ubiquitously expressing B6.C-Tg(CMV-cre)1Cgn/J mouse model (Jackson Laboratory, Bar Harbor, ME, USA, Jax Stock #006054) to generate global Gpr160 knockout animals. Animals homozygous for the Gpr160-floxed allele and hemizygous for CMV-Cre allele were used as experimental animals, whereas Gpr160-floxed littermates negative for the CMV-Cre allele were used as control animals. The Gpr160 knockout allele was confirmed in experimental animals through a multiplex PCR assay with PCR primers mGpr160-F1 (5′-GTCAACTAGGAATGCTTCCCTCCAC-3′), mGpr160-F2 (5′-GGACAGGAACTGAAGCTCGAGTC-3′), and mGpr160-R2 (5′-TCCACAATGTTGTCGAGCAGGC-3′). A 120 bp amplicon is detected in floxed samples, whereas a 162 bp amplicon is detected in Cre-mediated recombined samples with the Gpr160 coding region deleted (Fig. 1B and supplemental Figure 1, available at https://links.lww.com/PAIN/B974).
 Figure 1.:
Figure 1.: Generation of Gpr160-floxed and Gpr160 knockout mice. (A) CRISPR-Cas9 genome editing was used to introduce loxP sites flanking exon 4 of mouse Gpr160. (B) Genotypic analysis showed PCR amplicons consistent with expected sized corresponding to WT, floxed (FL), and knockout (KO) alleles. (C) No differences in weight gain were observed between FL and Gpr160 KO female or male mice at 2 and 5 months (P > 0.9; n = 7 FL males at 2 months and n = 8 FL males at 5 months; n = 4 Gpr160 KO males at 2 months and n = 5 Gpr160 KO males at 5 months; n = 3 FL females at 2 months and n = 7 FL females at 5 months; and n = 7 Gpr160 KO females at 2 months and n = 4 Gpr160 KO females at 5 months). Data are expressed as mean ± SD and analyzed by 3-way ANOVA with the Bonferroni post hoc test (C). ANOVA, analysis of variance; Gpr160, G-protein-coupled receptor 160; PCR, polymerase chain reaction. CRISPR-Cas9, Clustered regularly interspaced palindromic repeats-CRISPR-associated protein 9; loxP, locus of x-over P1; WT, wild-type.
2.2. Chronic constriction injury modelThe chronic constriction injury (CCI) model was performed using the well-established Bennett model.5 In brief, mice were put under general anesthesia (3% isoflurane/100% O2 inhalation and maintained on 2% isoflurane/100% O2 inhalation for the duration of surgery) and the left thigh was shaved. Nolvasan was applied before an incision (1-1.5 cm in length) was made in the middle of the lateral aspect. The sciatic nerve was exposed, and 6-0 silk sutures were ligated at 2 distinct spots (1 mm apart). Two ligations with silk thread are enough to recapitulate the pain-like behavior in mice, and a recent study came out showing that the number of ligations does not affect pain-like behavior, pathogenesis, and neuronal activation.32
2.3. Intrathecal injectionsIntrathecal (i.th.) injections were performed as previously described using the Wilcox method.36 The mice were placed under general anesthesia (3% isoflurane/100% O2 inhalation and maintained on 2% isoflurane/100% O2 inhalation), and the fur at the posterior end of the animal was shaved. A Hamilton syringe (Hamilton, Reno, NV) attached to a 30-G needle was inserted between the L5 and L6 vertebrae of the mice. Proper placement of the needle was confirmed by a reflexive tail flick or reflex of the hind limb. Five microliters (μL) of vehicle or test substance (s) was injected.
2.4. Behavioral testingGpr160 KO mice and their floxed control mice were behaviorally phenotyped to determine whether there were any strain differences in learning and memory, executive function, anxiety, depression, dexterity, and strength. Mechanical allodynia and cold allodynia were measured in all time course studies. All mice were weighed before being placed on an elevated wire-mesh platform in individual Plexiglas containers and allowed to acclimate for 0.5 to 1 hour. When mechanical and cold allodynia were assessed, mechanical allodynia was measured first, followed by a 15-minute break to prevent sensitization, then cold allodynia was measured.
2.4.1. Mechanical allodyniaMechanical allodynia was measured in mice using the Dixon up-and-down method as described previously.19 Calibrated von Frey filaments beginning at 0.16 g were placed on the plantar surface of the hind paw. Filament size increased until a positive response was observed such as clear paw withdrawal or shaking. A filament of 2.00 g was used as a cut-off and was given a positive response at 2.00 g. At least 4 readings were obtained after the initial positive response. The pattern of response was converted to a paw withdrawal threshold (PWT) at 50% using a method described previously.11 Mechanical allodynia was defined as a significant (P < 0.05) reduction in the PWT compared with the PWT of the baseline measurement.
2.4.2. Cold allodyniaCold allodynia was used to measure a cold innocuous stimulus as described previously.8,71 In brief, a small drop of acetone was placed on the plantar surface of the hind paw of the mice. The animal's response was monitored for 1 minute after application and scored based on a 4-point scale: 0, no response; 1, quick withdrawal or flick of the paw; 2, prolonged withdrawal of the paw or repeated flicking; and 3, repeated paw flicking and licking of the paw. To prevent oversensitization, the acetone was applied a total of 3 times with a 5-minute interval between each application. The response score was expressed as a % maximal response score [(the sum of the score from each trial/the sum of the maximum score) × 100].
2.4.3. Hot plateHot plate was used to test normal nociception. Mice were placed on the hot-plate set at 50°C, 52°C, and 55°C, starting at the lowest temperature. To prevent habituation, a 1-hour interval time was given between each temperature. Paw licking, shaking, or jumping was denoted as a response. The cutoff time was 20 seconds to minimize any tissue damage.
2.4.4. Tail flickTail flick assay was used to test acute thermal nociceptive responses that measures withdrawal latency of the tail from a noxious radiant heat source (model 37360; Ugo Basile, Gemonio VA, Italy) as described previously.16,20 The heat intensity was set to infrared, 50 and 60. Cutoff latencies were 10 seconds to prevent tissue damage. Data are reported as tail-flick latency (s).
2.4.5. T mazeThe aversive T maze is both a learning task based on working memory and a reference memory task.25 The T maze consisted of a black plastic alley with a start box at one end and 2 goal boxes at the other. The start box was separated from the alley by a plastic guillotine door that prevented movement down the alley until raised during training trials. An electrifiable floor of stainless-steel rods ran throughout the maze to deliver a mild, scrambled foot shock. Mice were not permitted to explore the maze before training. A block of training trials began when a mouse was placed into the start box. The guillotine door was raised, and a cue buzzer sounded simultaneously; 5 seconds later, foot shock was applied. The arm of the maze entered on the first trial was designated “incorrect” and the mild foot shock was continued until the mouse entered the other goal box, which in all subsequent trials was designated as “correct” for that mouse. At the end of each trial, the mouse was returned to its home cage until the next trial. Each mouse was trained until he made 1 avoidance. Training used an intertrial interval of 35 seconds, the buzzer was of the doorbell type sounded at 55 dB, and shock was set at 0.35 mA (Coulbourn Instruments scrambled grid floor shocker model E13-08). Retention was tested 1 week later by continuing training until mice reached the criterion of 5 avoidances in 6 consecutive trials. The results were reported as the number of trials to reach criterion for the retention test.
2.4.6. Y mazeThe Y maze test for spatial working memory was performed in a maze consisting of 3 arms (35 cm length × 5 cm width × 15.5 cm height per arm) at 120-degree angles as described.13 Each mouse was individually placed in 1 arm and allowed to freely explore the Y maze for 5 minutes. The percentage of perfect alternations (entry into an arm that differs from the previous 2 entries) over total arm entries was calculated as the ratio of the number of perfect alternations to the total number of possible alternations.
2.4.7. Novel object-place recognitionThe novel object-place recognition test (NOPRT) is a declarative memory task that involves the hippocampus when the retention interval was 24 hours and the prefrontal cortex when the retention interval is 1 hour after the initial exposure to the objects.34,53 During the training session, mice were exposed to 2 similar objects and allowed to explore for 5 minutes while an ANY-maze tracker (San Diego Instruments, San Diego, CA) recorded time spent investigating each object. The apparatus and objects were cleaned between each mouse. Twenty-four hours later, mice were exposed to one of the original objects and a novel object. The time spent investigating each object was again recorded. The novel object was made from the same material as the original object and approximately the same size but had a different shape. Each mouse received a 24 hours and a 1-hour test with different object used for each test. The test is based on the tendency of mice to spend more time exploring new objects than familiar ones. Thus, the more time spent with the novel object, the greater the retention or memory of the familiar object at 24 hours, as defined by the discrimination index (DI): DI= (time spent with novel object − time spent with familiar object)/(total time spent with both objects).
2.4.8. Puzzle boxThe puzzle box was used to test memory and complex problem solving. The test is performed over 4 consecutive days where each day the mice is challenged with obstacles that increase in difficulty as described previously.64 The box (61 cm × 30 cm) contains a barrier that has a large bright chamber and a small dark chamber. Animals are placed in the large bright chamber as the start and the goal is to solve the puzzle to enter the dark chamber in a limited amount of time (4 minutes/trial). The paradigm includes the following: D1 (trials 1-3), animals access the dark chamber through a tunnel where the entrance is completely opened; D2 (trials 4-6), the open tunnel is partially blocked by a guillotine; D3 (trials 7-9), the open tunnel is blocked by nesting paper; and D4 (trials 10-11), the open tunnel is blocked by a T-shaped cardboard plug. Mice that took more than 4 minutes to solve the first 3 trials on D1 of the puzzle were excluded from data analysis.
2.4.9. Elevated plus mazeTo further test for anxiety as a factor affecting performance behavioral tests, mice were subjected to an elevated plus maze test. Mice were placed onto the center zone of an elevated plus maze apparatus consisting of 4 perpendicular arms in the shape of a plus sign, elevated 50 cm above the floor. Each arm is 35.5 cm in length; 2 opposite arms are enclosed and 2 are open. The test consisted of placing a mouse in the central platform facing an enclosed arm and allowing it to freely explore for 5 minutes. The test arena was wiped with a damp cloth after each trial. The number of entries into the open and closed arms and the time spent in the open arms was recorded by an ANY-maze tracker. Anxiolysis is indicated by increased time spent in open arms and number of open arm entries.
2.4.10. Forced swim testDepression-like behavior was quantified as increased immobility in the forced swim test (FST). Mice were placed in an inescapable transparent tank (30 cm height × 20 cm diameters) with a water level of 15 cm and a temperature of 25 ± 1°C, and immobility time was recorded for the last 4 minutes of a 6-minute trial.73 A video recorder was placed above the maze to record the movement of the mouse. The greater the amount of time spent immobile indicates increased depressive behavior. After each trial, each mouse was placed in a cage with a warmed absorbent surface under a warming lamp until dry. During the test the mouse was continually monitored and rescued from drowning should it stop swimming or become compromised.
2.4.11. Strength testing 2.4.11.1. Grip strengthForelimb grip strength was measured by allowing the mouse to grasp a wire mesh connected to a grip strength meter (Columbus Instruments, Columbus, OH). The values were corrected by dividing the values for strength meter by body weight in kilograms.
2.4.11.2. Wire cage hangThe mouse was allowed to grip the top of a wire-mesh cage, which was then inverted 12 inches over a three-fourth-inch foam pad. The time the mouse was able to hang was recorded, and the test ended after a maximum of 3 minutes.
2.4.12. Parallel rod or foot slip testThe parallel rod or foot slip test is designed for assessing the effects of manipulations, brain damage and disease on the motor coordination, gait, and level of fatigue in mice. The testing arena consists of a 20 × 20-cm box with a grid floor positioned over a solid metal floor. The grid floor does not provide a shock. A mouse was placed in the arena and allowed to roam the box for 5 minutes. The apparatus has a computerized tracking system with an overhead camera that tracked the mouse and calculated the animal's speed, distance travelled, and foot slips, i.e., how many times a paw slips off the grid floor and touches the metal plate below (Stoelting, Wood Dale, IL). The test allows for the simultaneous measurement of ataxia and locomotor activity. An ataxia score was calculated using the ratio of foot slips per cm traveled. After the 5-minute test period, the mouse was returned to its home cage. The apparatus was cleaned and sanitized between mice.
2.4.13. Open fieldTo eliminate changes in locomotor activity and anxiety as a factor affecting performance on subsequent behavioral tests, mice were subjected to an open field test. Mice were placed in an open field (67 cm × 57 cm × 24 cm) for 5 minutes. Distance traveled, time spent in center and peripheral zones, and number of entries into the center zone (33.5 cm × 28.5 cm) of the open field were recorded by an ANY-maze tracker.
2.5. Estrous smearsFive to seven days before and after the experiment, vaginal smears were obtained from female mice to ensure normal cycling as described previously.8,9 Smears were taken in the morning to maintain consistency. Approximately 10 µL of sterile saline was used to collect cells and they were placed on a glass slide. Cells were read fresh under a light microscope, and the stage of the estrous cycle was determined by the proportion of cell types. Females cycled normally and treatment did not alter the estrous cycle.
2.6. Compounds and administrationCompounds were given either intraplantar (i.pl.) or intrathecally (i.th.). All i.pl. injections were given in a final volume of 20 µL, and all i.th. injections were given in a final volume of 5 µL.
Cocaine- and amphetamine-regulated transcript peptide (55-102) (#003-62; purity ≥ 95%) from Phoenix Pharmaceuticals was reconstituted in distilled water. When CARTp was given i.th., it was diluted in artificial cerebral spinal fluid (aCSF; LRE-S-LSG-1000-1; EcocyteShop, Austin, TX). When CARTp was given i.pl., it was diluted in sterile saline. In all studies with CARTp, the dose given was 30 ng, a previous dose known to cause sensitivity.75
In pertussis toxin (PTX) studies, 0.1 µg of PTX (Sigma-Aldrich # P2980-50 µg) or Veh (5% glycerol in aCSF when given i.th. or 5% glycerol in saline when given i.pl.) was given 3 days before CARTp administration as previously described.7,63 BL (D1) was assessed before PTX and before CARTp injection, on D3, to ensure injections did not cause sensitivity.
2.7. StatisticsAll data were analyzed by GraphPad Prism (version 8.00 for Windows, GraphPad Software, San Diego CA) and are expressed as mean ± SD or SEM of n number of animals. Cold allodynia and log10-transformed von Frey behavioral data were analyzed by 2-tailed, 2-way analysis of variance (ANOVA) with the Bonferroni post hoc test. Weight gain and hot-plate test data were analyzed by 3-way ANOVA with the Bonferroni post hoc test. Cognitive behavioral results were analyzed by 2-tailed, 1-way or 2-way ANOVA with the Bonferroni post hoc analysis as noted. Outliers, defined as values that were more than 2 standard deviations from the group mean, were removed from the analysis. One floxed (FL) female was excluded from the open-field test as an outlier. A P value ≤ 0.05 was considered significant for all behavioral analysis.
3. Results 3.1. Generation of Gpr160 global KO miceTo further investigate the contribution of GPR160, we used CRISPR-Cas9 technology to generate Gpr160-floxed (FL) mice (Fig. 1A). These Gpr160-floxed mice were then crossed with a ubiquitously expressing CMV-Cre mouse model to produce the Gpr160 global knockout (KO) mice. The Gpr160 knockout allele was first confirmed through PCR of DNA extracted from tail biopsies (Fig. 1B) and further confirmed through PCR of DNA extracted from the spinal cord (Supplementary Figure 1B, available at https://links.lww.com/PAIN/B974). Polymerase chain reaction amplicons generated by using primers flanking exon 4 corresponded to the expected KO allele length with the coding region of exon 4 removed (162bp). Overall, the Gpr160 KO mice are fertile and healthy with no noticeable physical abnormalities. There were no differences in weights between Gpr160-floxed mice and Gpr160 KO mice in either sex even after 5 months of age when fed a chow diet (Fig. 1C).
3.2. Spinal cocaine- and amphetamine-regulated transcript peptide/G-protein-coupled receptor 160 induces behavioral hypersensitivitiesIntrathecal (i.th.) injections of CARTp evoked a robust time-dependent and GPR160-dependent development of mechanical and cold allodynia (behavioral hypersensitivities) in Gpr160-floxed male and female mice which were almost completely blocked in the Gpr160 KO mice (Figs. 2A and B). Studies have shown CARTp signals using Gi/o-GPCR.46,63 Pretreatment with pertussis toxin (PTX, an ADP-ribosylating toxin that inactivates Gi/o proteins) given i.th. blocked CARTp-induced mechanical and cold allodynia in male and female mice, further supporting Gi/o-coupled signaling as a mode of action (Figs. 2C and D).7,47 Results shown in Figure 2 include a mixed population of male and female mice because major sex-dependent differences were not observed when analyzed separately (Supplemental Figure 2A-H, available at https://links.lww.com/PAIN/B974).
 Figure 2.:
Figure 2.: CARTp administration in the spinal cord causes behavioral hypersensitivities through GPR160 in a Gi/o-dependent manner. (A and B) A single i.th. injection of CARTp (30 ng) induced profound mechanical (A) and cold allodynia (B) in Gpr160 FL/FL (n = 7 males and n = 5 females) compared with Gpr160 KO/KO (n = 7 males and n = 6 females) mice. (C and D) CARTp-induced mechanical (C, n = 6 males and n = 4 females per group) and cold (D, n = 4 males and n = 4 females per group) allodynia were prevented in ICR mice that received an i.th. injection of PTX (0.1 µg) 3 days before CARTp administration. Data are expressed as mean ± SD and analyzed by 2-tailed, 2-way repeated measures ANOVA with the Bonferroni post hoc test (A–D). *P < 0.05 vs 0 hours or D3, †P < 0.05 vs Veh + CARTp and #P < 0.05 vs Gpr160 FL/FL. ANOVA, analysis of variance; CARTp, cocaine- and amphetamine-regulated transcript peptide; FL, floxed; GPR160, G-protein-coupled receptor 160; KO, knockout; PTX, pertussis toxin ICR, Institute of Cancer Research.
3.3. Peripheral cocaine- and amphetamine-regulated transcript peptide/G-protein-coupled receptor 160 induces behavioral hypersensitivitiesThe role of CARTp in peripheral sensitization is not known. Intraplantar (i.pl.) injection of CARTp in male and female mice evoked time-dependent behavioral hypersensitivities that were blocked by i.pl. injections of PTX supporting Gi/o-coupled signaling as a mode of action in the spinal cord and in the periphery (Figs. 3A–D).47 No effects were observed in the contralateral paws (data not shown). Using a calibrated caliber, paw thickness (an index of paw edema) was measured and was not different between the vehicle and CARTp groups, suggesting that CARTp injection does not evoke a measurable inflammatory response (data not shown). I.pl. injection of CARTp in Gpr160 KO mice failed to evoke behavioral hypersensitivities establishing GPR160 dependency of the response (Figs. 3E and F). Results shown in Figure 3 include a mixed population of male and female mice because major sex-dependent differences were not observed when analyzed separately (Supplemental Figure 3A-H, available at https://links.lww.com/PAIN/B974).
 Figure 3.:
Figure 3.: CARTp causes behavioral hypersensitivities in the periphery through GPR160 in a Gi/o-dependent manner. (A and B) I.pl. injection of CARTp (30 ng) induced profound mechanical (A, males: n = 7 Veh-treated group and n = 8 CARTp-treated group; n = 4 females per group) and cold (B, n = 5 males and n = 4 females per group) allodynia in ICR mice. (C and D) CARTp-induced mechanical (C) and cold allodynia (D) were reversed in ICR mice that received an i.pl. injection of PTX (0.1 µg) 3 days before CARTp administration (n = 4 males per group). (E and F) CARTp-induced mechanical (E) and cold allodynia (F) were lost in Gpr160 KO/KO mice (n = 5 males and n = 6 females) when compared to Gpr160 FL/FL mice (n = 5 males and n = 5 females). Data are expressed as mean ± SD and analyzed by 2-tailed, 2-way repeated-measures ANOVA with the Bonferroni post hoc test (A–F). *P < 0.05 vs 0 hour or D3, †P < 0.05 vs CARTp or Veh + CARTp, #P < 0.05 vs Gpr160 FL/FL. ANOVA, analysis of variance; CARTp, cocaine- and amphetamine-regulated transcript peptide; FL, floxed; GPR160, G-protein-coupled receptor 160; KO, knockout; PTX, pertussis toxin I.pl., intraplantar; ICR, Institute of Cancer Research.
3.4. Nociceptive behaviors in naïve and nerve-injured miceChronic constriction of the sciatic nerve in male and female mice lead to a time-dependent development of behavioral hypersensitivities in Gpr160-floxed mice but not in age-matched and sex-matched Gpr160 KO, suggesting that GPR160 is necessary for the development of neuropathic pain (Figs. 4A and B). These data support our previous findings that using genetic and immunopharmacological approaches to block GPR160 attenuated and reversed behavioral hypersensitivities in 2 different animal models of neuropathic pain.75 Major sex differences in mechanical or cold allodynia were not observed when analyzed separately (Supplemental Figure 4A–D, available at https://links.lww.com/PAIN/B974). By contrast, responses to the hot-plate test and tail-flick latency were similar in naïve Gpr160 KO and Gpr160-floxed mice (Figs. 4C and D). These results support our previous findings that blocking GPR160 does not alter normal thermal nociceptive responses.75
 Figure 4.:
Figure 4.: Mice lacking GPR160 do not develop CCI-induced hypersensitivities. (A and B) Gpr160 KO/KO mice (n = 3 males and n = 6 females) did not develop CCI-induced mechanical (A) and cold (B) allodynia in the ipsilateral paw compared with the Gpr160 FL mice (n = 5 males and n = 5 females) after 21 days. (C) Withdrawal latency was not different between Gpr160 FL/FL mice and Gpr160 KO/KO within the same sex (P = 0.17; n = 4 males and 3 females per group). (D) Tail-flick latency was not different between Gpr160 FL/FL and Gpr160 KO/KO male mice at 50 or 60 infrared intensity (P = 0.71; n = 6 per group). Data are expressed as mean ± SD and analyzed by 2-tailed, 2-way repeated measures ANOVA with the Bonferroni post hoc test (A, B, D) or 3-way ANOVA with the Bonferroni post hoc test (C). *P < 0.05 vs D0 and #P < 0.05 vs Gpr160 FL/FL. ANOVA, analysis of variance; CCI, chronic constriction injury; FL, floxed; GPR160, G-protein-coupled receptor 160; KO, knockout.
3.5. Pharmacologic characterization of G-protein-coupled receptor 160 KO mice on cognitive behaviorsThere were no differences between males and females of each strain in T-maze long-term spatial memory, Y-maze working spatial memory, NOPRT 1-hour short-term recognition memory, and NOPRT 24-hour long-term recognition memory (Figs. 5A–D). In the puzzle box test of complex problem solving and memory, there were no differences in problem solving ability in the female Gpr160 KO mice compared with their age-matched cohort (Fig. 5F). Similarly, there were no differences in puzzle box performance in male Gpr160 KO mice compared with the male Gpr160-floxed cohort, except for trial 9 where the Gpr160 KO mice spent less time solving the puzzle task (Fig. 5E). In the elevated plus maze test of anxiety, there was no difference in time spent in open arms between male Gpr160-floxed and Gpr160 KO mice; although there was a slight increase in time spent in open arms (indicating they are less anxious) in the female Gpr160 KO mice compared with Gpr160-floxed cohort, it was not statistically significant (Fig. 5G). Although there were no differences in the forced swim test of depression in the male Gpr160 KO mice compared with their control cohort, the female Gpr160 KO mice spent significantly more time immobile than the female control mice, suggesting an increase in depression-like behavior (Fig. 5H). In the parallel rod test of ataxia, the Gpr160 KO female mice had more foot faults (slips) than their control female cohort, but there was no difference between the male mice (Fig. 5I). However, there was no difference in the impairment ratio in either sex or genotype, suggesting there are no motor impairments (Fig. 5I). There were no differences in forelimb grip strength between the male Gpr160 KO and Gpr160-floxed mice; however, the Gpr160 female KO mice had a reduced forelimb grip strength compared with their female control cohort (Fig. 5J). However, in the wire hang test of strength, there were no differences in the ability to hang upside down between the Gpr160-floxed and Gpr160 male or female mice (Fig. 5K). In the open-field measure of general activity, there were no differences in the total distance traveled in either the male or female mice of either genotype (Fig. 5L).
 Figure 5.:
Figure 5.: Phenotypic characterization of the Gpr160 KO mice. (A–D) Sex and genotype (FL vs Gpr160 KO) of the mice had no significant effect on working, spatial, short-term, or long-term memory behaviors in the T maze (A; P = 0.28), Y maze (B; P = 0.49), NOPRT 1 hour (C; P = 0.86), and NOPRT 24 hours (D; P = 0.20) (FL: n = 8 males and 10 females; Gpr160 KO: n = 5 males and n = 6 females). (E) Loss of GPR160 in male mice had no significant impact on their performance in the puzzle box except during trial 9, where the male knockout mice performed better (FL: n = 8; Gpr160 KO: n = 5). (F) the loss of GPR160 in female mice had no significant impact on their performance in any puzzle box trials (P = 0.25, FL: n = 10; Gpr160 KO: n = 6 per group). (G) Sex and genotype (FL vs Gpr160 KO) of the mice did not significantly alter anxiety-like behavior in the elevated plus maze (P = 0.11; FL: n = 7 males and n = 9 females; Gpr160 KO: n = 4 males and n = 5 females). (H) In the forced swim test of depressive-like behavior, Gpr160 KO female mice spent significantly more time immobile than the FL female cohort (FL: n = 10; Gpr160 KO: n = 6). There were no significant differences in the male behavior (P > 0.99; FL: n = 8; Gpr160 KO: n = 5). (I) In the parallel rod test of ataxia, female (n = 6), but not male (n = 5), Gpr160 KO mice had more foot slips on the rods compared with their FL cohort (n = 9 females and n = 8 males). There was no difference in the impairment ratio for male and female mice. (J) In the measure of strength, female Gpr160 KO (n = 5), but not male, Gpr160 KO (n = 5) mice had a reduction in front paw grip strength compared with their FL cohort (n = 8 males and 10 females). (K) There was no difference in the ability to hang upside down in the cage between FL and Gpr160 KO male (P > 0.99; FL: n = 8; Gpr160 KO: n = 5) or female mice (P = 0.82; FL: n = 10; Gpr160 KO: n = 6). (L) There was no difference in the general activity in male (P = 0.07; FL: n = 8; Gpr160 KO: n = 5) or female mice (P > 0.99; FL: n = 9; Gpr160 KO: n = 6). Data are expressed as mean ± SD and analyzed by 1-way ANOVA with the Bonferroni post hoc test (A–D, G–L) or 2-tailed, 2-way repeated-measures ANOVA with the Bonferroni post hoc (E, F). *P < 0.05 vs FL. ANOVA, analysis of variance; FL, floxed; GPR160, G-protein-coupled receptor 160; KO, knockout; NOPRT, novel object-place recognition test.
4. DiscussionCurrently, there are no known small-molecule antagonists for GPR160 and there are limitations to studying the physiological role and disease states of CARTp/GPR160 by using genetic (eg, siRNA) and immunological approaches (eg, neutralizing antibody). This study uses newly generated Gpr160 global KO mice to test the importance of GPR160's role in CARTp-induced behavioral hypersensitivities, nerve injury–induced neuropathic pain mouse model, and affective behaviors associated with psychological comorbidities prevalent in patients with chronic pain.
Gpr160 KO mice are overall healthy and fertile. No obvious physical abnormalities were observed. Although CARTp is a widely known anorectic peptide, only recently was GPR160 shown to be involved in feeding and required for CARTp's anorexigenic behavior.33,59,61 Compared with age-matched controls, the Gpr160 KO mice did not differ in body weight in either sex up to 5 months after birth when fed a chow diet. Different strains of Cartp KO mice also showed no body weight changes at a young age compared with controls when fed a chow diet.2,5
留言 (0)