
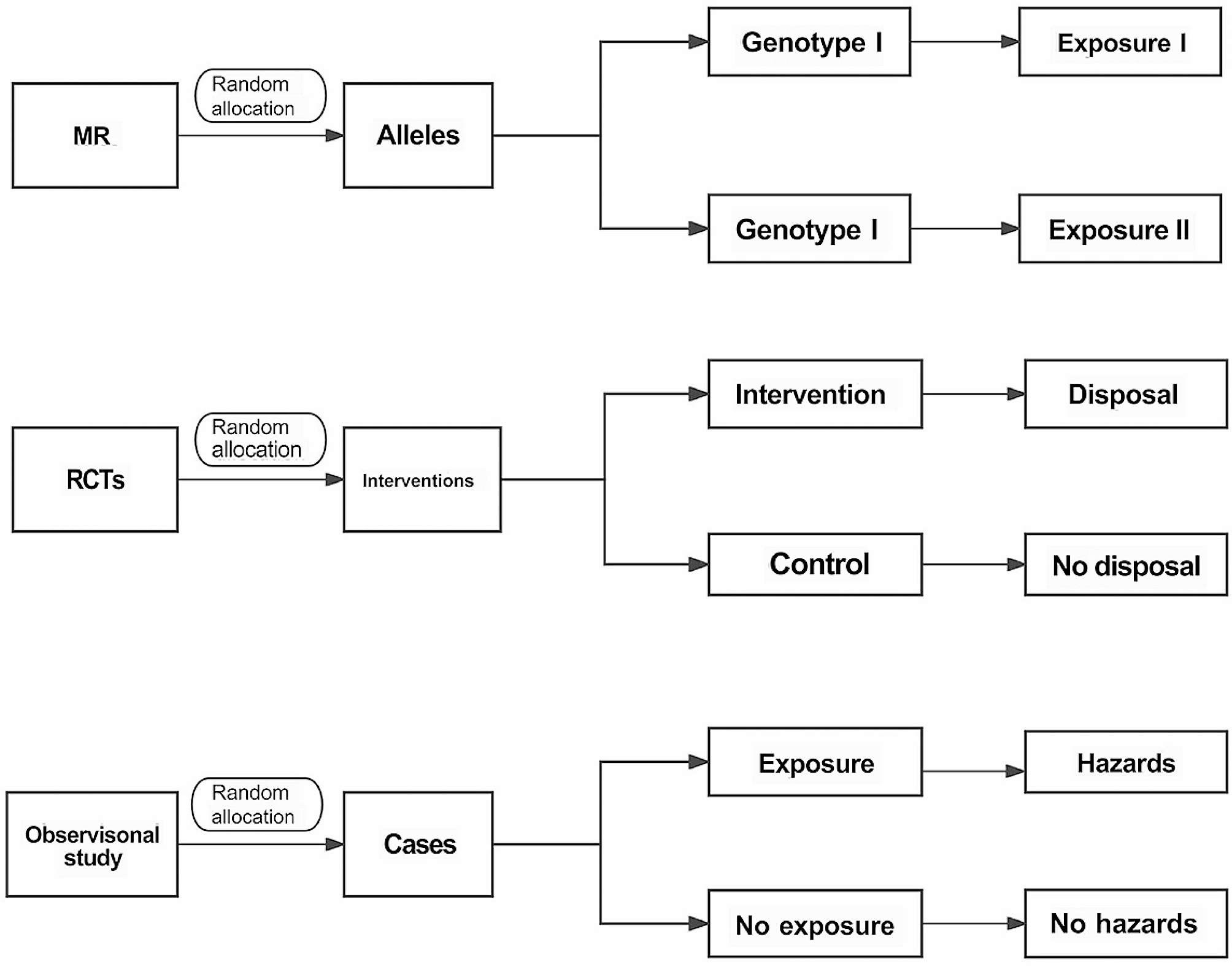
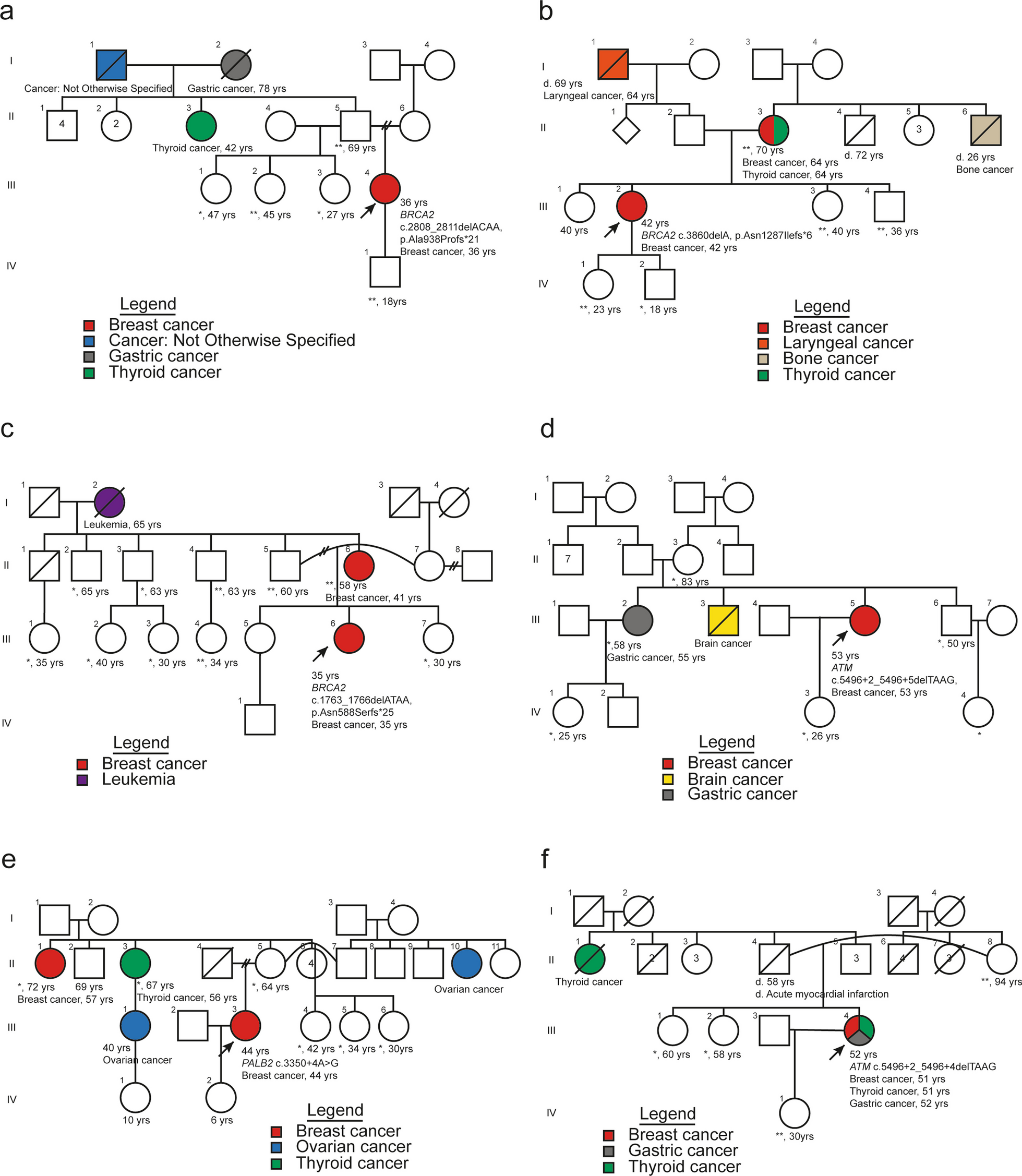
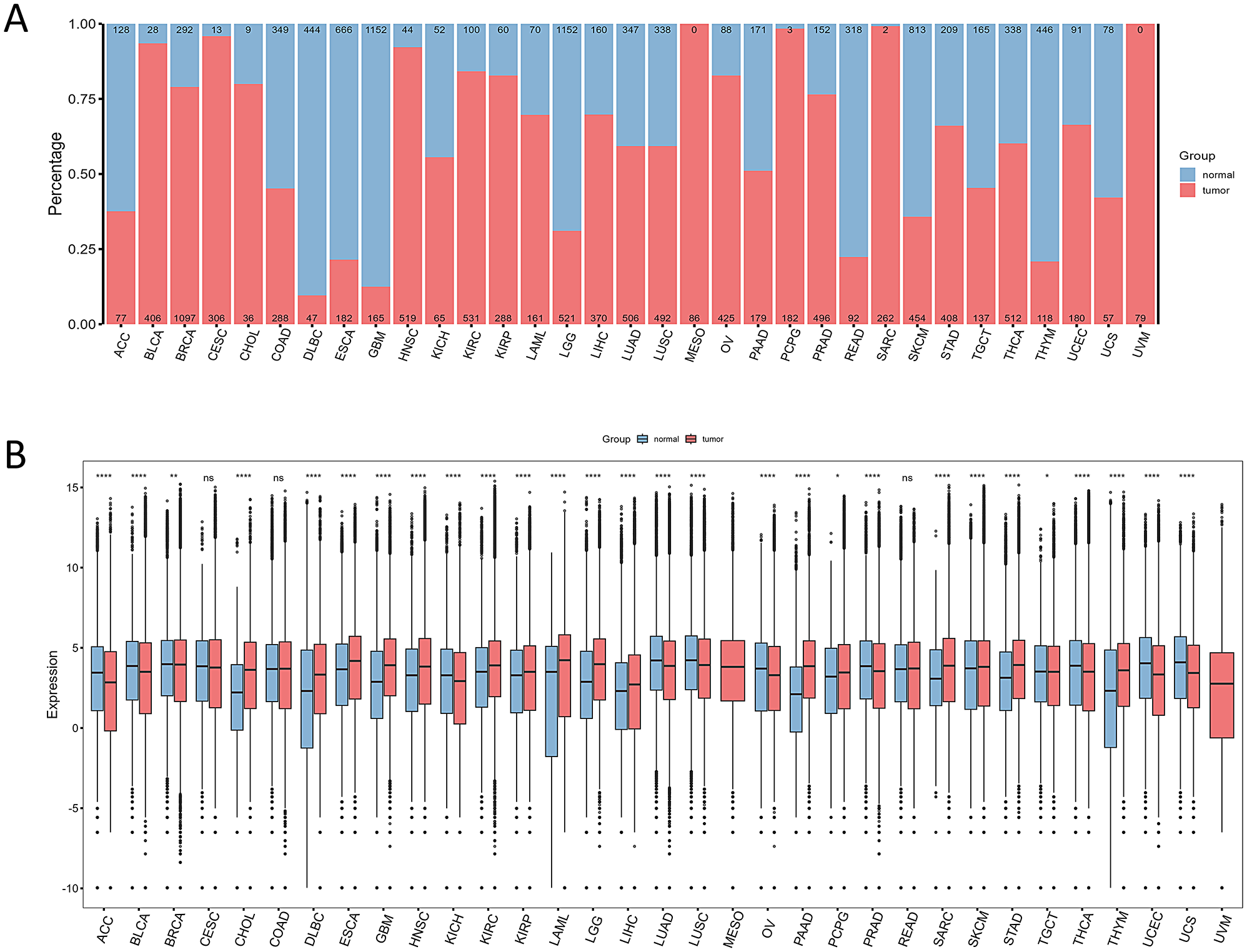
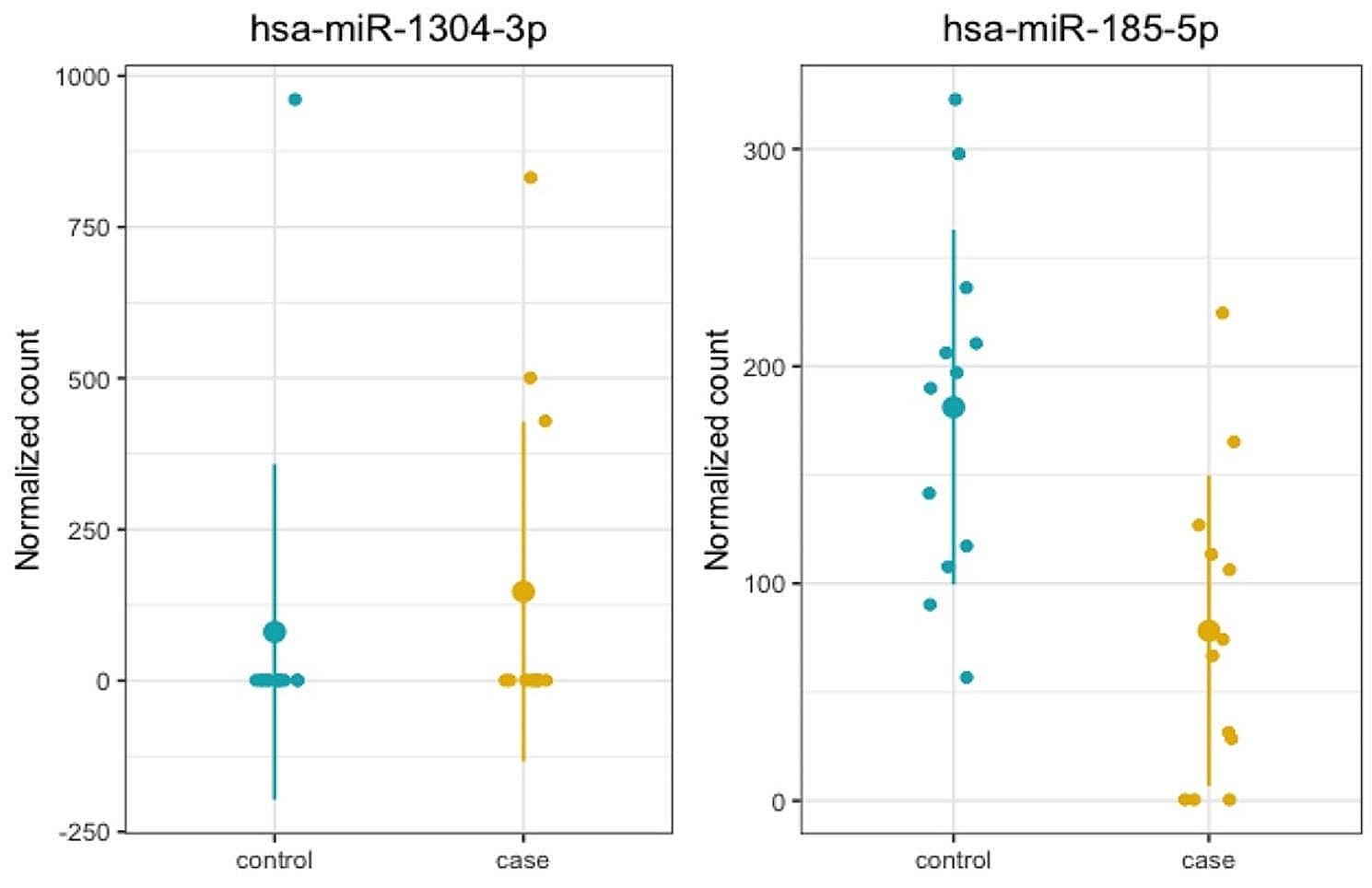
記住我

A study cohort of 51 individuals of Southeast Asian ethnicity (36 CI patients plus 15 age-matched healthy subjects) were assessed clinically and further analyzed by microbiome sequencing, whole exome sequencing (WES), and immunophenotyping (Fig. 1A). First, we examined the clinical patterns of each disorder and compared them to related genetic mutations as defined and categorized by expert dermatologists, microbiologists, and geneticists in Vietnam. We categorized the CI patients (n = 36) into having one of 7 CI disorders: IV (n = 15), HI (n = 8), LI (n = 3), EI (n = 4), Trichothiodystrophy (TTD, n = 3), Arthrogryposis renal dysfunction cholestasis (ARC, n = 1), and Sjögren-Larsson Syndrome (SLS, n = 2) and compared them alongside healthy age-matched subjects (n = 15). The median duration of follow-up was 4.3 years with patient age (average 10.2 years old) ranging from neonatal (birth) to adults (40 years). Clinical phenotypes of some of the CI patients are shown (Fig. 1B).
Fig. 1
Study design and clinical phenotypes found in Southeast Asian individuals with congenital ichthyosis (CI). A Schematic overview of study design and methods used. B Clinical photographs of representative individuals with harlequin ichthyosis (HI1, HI5, HI7, HI8), lamellar ichthyosis (LI1), epidermolytic ichthyosis (EI1), trichothiodystrophy/XPD (TTD1), ichthyosis vulgaris (IV1), arthrogryposis, renal dysfunction, and cholestasis (ARC1) syndrome
To characterize the frequency of specific clinical manifestations in CI, our cohort was evaluated for 10 findings: hyperkeratosis, itch, microbial infection, erythroderma, respiratory difficulties, sepsis, collodion membrane at birth, loss of heat, blistering, and cardiac abnormalities. Hyperkeratosis and itch were the most common clinical manifestations of CI (100% and 97.2%, respectively), followed by microbial infection (94.4%) and erythroderma (72.2%) (Table 1). 52.78% of CI patients demonstrated respiratory comorbidities and/or sepsis. There were 6 lethal cases due to infant sepsis (4 HI, 1 LI, and 1 EI). Ten other cases of childhood sepsis were evident in HI (3 out of 8) or in IV (7 out of 15) but were treatable with antibiotics (Additional file 14: Table S1 and Additional file 15: Table S2). Most HI and LI cases also presented with neonatal-onset chronic diarrhea, episodes of fever, recurrent pyodermas, oral candidiasis, and otitis externa. Aside from sepsis, one TTD patient developed necrotizing fasciitis of the nose following Burkholderia pseudomallei infection. Interestingly, 19/36 CI patients (53%) exhibited some form of mental health difficulty (Additional file 14: Table S1).
Table 1 Clinical phenotypes of congenital ichthyosis (CI) cohortNext, we performed WES and genetic linkage analysis in the 36 CI individuals. 31 pathogenic/likely pathogenic mutations (based on American College Medical Genetics) and 20 novel variants were identified (Table 2). Four novel variants were found in more than one CI patient in our cohort. We identified compound heterozygous and homozygous segregated mutations linked to autosomal recessive loss-of-function of ABCA12, TGM1, ALDH3A2, and ERCC2. The genetic mutations are mapped onto their corresponding protein domain (Fig. 2) and where possible, we analyzed the protein structure alterations caused by the CI-related missense mutations (Additional file 2: Fig. S1, Additional file 3: Fig. S2, Additional file 4: Fig. S3, Additional file 5: Fig. S4).
Table 2 Whole exome sequencing (WES) analysis of 36 CI patientsFig. 2
Pathogenic variants identified in cohort of Southeast Asian CI patients. Schematic representation of mutated gene domains with annotated missense, nonsense, splicing, and frameshift mutations identified in affected individuals with ABCA12, TGM1, KRT1, FLG, ERCC2, VSP33B, and ALDH3A2 mutations. “Coupling mutations” refers to variants identified together in the same patient. Variants highlighted with yellow experienced sepsis: we note that most of sepsis cases happened with double mutation, while 70% of the ABCA12 variants associated with sepsis in our cohort were adjacent to the ABC1 or ABC2 domains; KRT1 G488V variant associated with sepsis occurred at the C-terminus in the highly conserved TYR*LLEGE motif known to be critical for intermolecular and higher order filament interactions (see Additional file 3: Fig. S2); IV patients who experienced sepsis had either frameshift or nonsense mutations in the N-terminal half of FLG whereas those having mutations more C-terminal did not experience sepsis
To further evaluate our cohort for clinically relevant genotype–phenotype correlations, Fisher exact tests and odds ratio calculations were performed for 12 clinical findings over the 7 distinct CI genotypes (Additional file 18: Table S5). We observed statistically significant association of intensive care unit (ICU) admission with ABCA12 mutation, death examination with ABCA12 and TGM1 mutations, sepsis with ABCA12 mutation, respiratory problems with FLG mutation, collodion membrane at birth with ABCA12 and TGM1 mutation, blistering with ABCA12 and FLG mutation, dry and thick skin with FLG mutation, impaired wound healing with FLG mutation, itch with ABCA12 and FLG mutations, and bacterial infection with FLG mutation.
CI patient microbiome signaturesTo better understand the risk of infections in CI, we comprehensively examined the CI patient microbiome (n = 36) by taking superficial swabs of sebaceous skin (facial region), dry skin (olecranal or patella regions) and moist skin (antecubital or popliteal region) and compared the results to age-matched healthy subjects (n = 15; Fig. 1A). We focused on the 4 dominant phyla—Actinobacteria, Firmicutes, Proteobacteria, and Bacteroidetes. Actinobacteria and Firmicutes both showed decreased average relative abundance of colony forming units (cfu) in CI dry, moist, and sebaceous skin compared to the respective locations in healthy controls (Fig. 3A). Proteobacteria showed decreased relative abundance of cfu in dry CI skin compared to healthy controls, but increased abundance in moist and sebaceous skin. Bacteroidetes appeared stable to slightly increased in relative abundance of cfu across dry and sebaceous skin locations in CI patients compared to healthy controls, but slightly decreased in abundance in moist skin. Interestingly, Fungi were increased in sebaceous skin in CI patients compared to healthy controls.
Fig. 3
Microbiome profile in CI patients. A Bacterial community composition at phylum level (Actinobacteria, Firmicutes, Proteobacteria, Bacteroidetes, fungi, and other) represented as average relative abundance from healthy (left) and CI patients (right) and three different skin sites: dry, moist, and sebaceous. B Relative abundance of operational taxonomic units (OTUs) in healthy (left) and CI patients (right) from three different skin sites (dry, moist, and sebaceous) by White’s non-parametric t-test. Circle sizes are proportional to the number of sequences within for the phyla Actinobacteria, Firmicutes, Proteobacteria, Bacteroidetes, fungi, and gram positive/gram negative bacteria. C–M Abundance of specific bacterial species for each CI clinical type: C Cutibacterium acnes; D Lawsonella clevelandensis; E, Bacteroides uniformis; F Bacteroides vulgatus; G Staphylococcus capitis; H Staphylococcus aureus; I Staphylococcus hominis; J Staphylococcus epidermidis; K Moraxella osloensis; L Burkholderia pseudomallei; M Malassezia species. Data expressed as dots with different colors for dry (green), sebaceous (red), and moist (blue) skin with mean ± SD standard error. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001
Next, we compared the operational taxonomic units (OTU) sequence reads between the CI patients (n = 36) and healthy controls (n = 15) for dry, moist, and sebaceous skin locations (Fig. 3B). We observed reduced OTU counts of Actinobacteria, Firmicutes, Proteobacteria, and Gram-positive microbes across moist and sebaceous skin, but no significant difference in Bacteroidetes OTU counts (Fig. 3B). Analysis of the OTU sequences at the species level identified a significant reduction in species within the Actinobacteria, Firmicutes, and Bacteroidetes phyla for certain CI types (Fig. 3C–L). For Actinobacteria, CI patients generally had reduced levels of Cutibacterium acnes compared to healthy controls, with the reduction in C. acnes being statistically significant for IV, HI, LI, and EI types (Fig. 3C). L. clevelandensis demonstrated a mixed pattern, with significant elevations in IV patients but significant absence in LI and HI patients (Fig. 3D). For the Bacteroidetes phylum, Bacteroides uniformis was significantly elevated in EI and IV patients, and B. vulgatus had significant reductions in IV, HI, LI, EI, and TTD patients (Fig. 3E, F). In the Firmicutes phylum, Staphylococcus capitis, S. hominis, and S. epidermidis showed significantly reduced levels in all CI patients compared to healthy controls (Fig. 3G, I, J).
In contrast to the reductions in C. acnes, S. capitis, S. hominis, S. epidermidis, and B. vulgatus, S. aureus of the Firmicutes phylum showed statistically significant elevations in IV and EI patients versus healthy controls, and statistically non-significant elevations in dry skin of LI patients (Fig. 3H). Soil-dwelling and zoonotic disease-associated bacteria from the Proteobacteria phyla were also identified in certain CI patients. Burkholderia pseudomallei was exclusively increased in TTD patients (Fig. 3L), whereas Moraxella osloensis was increased in multiple CI types, most significantly IV, HI, EI, and TTD (Fig. 3K). Lastly, LI and HI were unique among the CI types in demonstrating a marked reduction in Malassezia species, in contrast to IV, EI, and TTD patients who experienced higher Malassezia abundance (Fig. 3M).
To evaluate the impact of CI genotype on the presence of various bacteria, we plotted bacterium abundance against CI gene mutation (Additional file 13: Fig. S12). For C. acnes, the difference in the amount of C. acnes between Healthy (WT) skin and skin with CI variants suggests that genotypes may influence the abundance of C. acnes. This may impact biological properties of the skin such as moisture level, pH, lipid barrier, or the skin's immune capabilities determined by the genotypes. Bacteroides vulgatus is part of the normal flora of the human skin, but was found in lower abundance in certain CI genotypes. The significant increase in S. aureus levels in individuals with FLG and KRT1 genotypes indicate that these genetic variants create favorable conditions for the proliferation of S. aureus on the skin. B. pseudomallei uniquely was elevated in ERCC2 genotypes compared to WT and other CI genotypes. Lastly, we observed a significant increase in Malassezia spp. between individuals with several genotypes versus those in the WT group.
Principal co-ordinate analysis identifies CI microbiome subgroupsTo investigate differences in pathogenic and commensal bacteria in healthy and CI patients across skin locations, the superficial swab OTU sequence data was subjected to principal co-ordinate analysis (PCoA). We identified dysbiosis clusters by collecting high-ratio non-pathogenic (principal component 1, PC1, x-axis) and high ratio pathogenic (principal component 2, PC2, y-axis) microbiomes observed in CI patients (Fig. 4A). Patients segregated into 5 distinct clusters following analysis of dry, moist, and sebaceous skin samples: cluster P1–healthy control subjects were high in PC1 and low in PC2; cluster P2–IV patients were high in PC2 and low in PC1 (FLG variants); cluster P3–EI and TTD patients were low in PC1 and variably elevated in PC2 in moist and sebaceous skin (KRT1 and ERCC2 variants, respectively); cluster P4–LI and HI patients were consistently lowest in both PC1 and PC2 across all skin sites (TGM1 and ABCA12 variants, respectively); and cluster P5–ARC and SLS patients were low in PC1 and variably elevated in PC2 in sebaceous skin only (VPS33B and ALDH3A2 variants, respectively) (Fig. 4B). Principal component analysis (PCA) validated the PCoA (Additional file 6: Fig. S5).
Fig. 4
Dysbiosis clustering of CI patients and the pathogenic microbiome in wounded skin of CI patients. A PCoA plots showing beta diversity metrics in dry, moist, sebaceous, and wounded CI skin, colored according to CI type. B CI microbiome dysbiosis clusters and distribution of P1-P5 in each skin region, including Dry, Sebaceous (Seb), Moist, and Wounded were determined by principal co-ordinate analysis in panel A (PC1/commensal, PC2/pathogenic). Specifically, the sample composition for the Dry, Seb, and Moist areas encompassed 7 Healthy, 3 TTD, 1 ARC, 2 SLS, 8 HI, 3 LI, 15 IV, and 4 EI samples. Meanwhile, the sample composition for the Wound area consisted of 1 Healthy, 2 TTD, 1 SLS, 6 HI, 3 LI, 9 IV, and 2 EI samples. C (top) Dynamic changes in abundance of S. aureus and S. epidermis communities in non-wounded (blue) and wounded (red) skin from representative CI patients (IV, EI, TTD, LI, HI, SLS) compared with healthy controls. (bottom) Heatmap diagram showing each CI group compared to healthy controls, and each group compared to each other, using two-way ANOVA analysis. Results of each bar are expressed as mean ± SEM standard error. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001. D, E After wounding CI patients and healthy controls with 5 mm punch biopsy, healing time, measured as the change in percent wound area over 7 days, was delayed for the 4 CI dysbiosis clusters P2-P5 compared to healthy P1 controls in the absence of antibiotics (D), but in the presence of β-lactam antibiotics healing time improved to levels comparable to healthy controls in (E). F, G Measurement of relative colonies of Staphylococcus aureus (F) or Staphylococcus epidermidis (G) across the 5 dysbiosis clusters in wounded skin over 36 h
Wound infection impairs healing of CI skinTo investigate how the microbiota changes in CI patients when their skin is wounded, we introduced 5 mm punch biopsy wounds in a CI wound-cohort (n = 32; the ARC patient from P5 did not participate) and matched healthy controls (n = 5) and evaluated them for microbial infection, wound healing time, and inflammation. Using PCoA on collected OTU sequences, we demonstrated that wounds in CI skin are characterized by a major shift in the dysbiosis clusters of CI patients to a very high PC2 and low PC1 state (Fig. 4A, far right panel). This same shift occurred for the healthy controls.
Next, we delineated the relative abundance of microbial species in CI wounds. In non-wounded skin (close to wound region), S. epidermidis abundance was high for healthy and CI skin, with IV and EI patients having the least abundant S. epidermidis (Fig. 4C, blue, center panel). In non-wounded skin, S. aureus abundance was much lower than S. epidermidis, though higher among CI types than healthy controls (Fig. 4C, blue, left panel). In contrast, wounded skin demonstrated a large shift towards more abundant S. aureus (mean = 48.5% total relative abundance) for healthy and CI patients, with concomitant reduction in S. epidermidis (mean = 43.76% total relative abundance) (Fig. 4C, red), as supported by two-way ANOVA analysis (Fig. 4C bottom). Streptococcus pyogenes (mean = 3.3% total relative abundance) and low proportions of the zoonotic pathogen Streptococcus suis (0.3% relative abundance) were also observed more in wounded skin compared to healthy control skin (Additional file 7: Fig. S6A).
Some Gram-negative, facultative anaerobes such as Pseudomonas aeruginosa and Campylobacter demonstrated different relative abundance among dysbiosis clusters. For example, P. aeruginosa (2.15% relative abundance) showed the greatest elevated abundance in P2 (IV) and P3 (EI/TTD), a smaller elevation in P5 (SLS), but was not detected in P4 (LI/HI), compared to healthy controls (Additional file 7: Fig. S6A). Campylobacter (0.73% relative abundance) similarly showed elevated abundance in P2, P3, and the LI patients in P4. Candida albicans (0.82% relative abundance) was similar in abundance across healthy controls and IV patients (P2), but showed increased abundance in EI patients (P3) and SLS patients (P5), and decreased abundance in P4 and among TTD patients (P3). Methicillin-resistant S. aureus (MRSA) was most abundant in HI patients (P4) (Additional file 7; Fig. S6A and Additional file 8: Fig. S7A), and also observed in IV (P2). Using real-time PCR, we observed elevated HPV virus (1.45% relative abundance) in TTD (P3) and LI (P4) patients (Additional file 8: Fig. S7B).
To further evaluate whether the abundance of pathogenic microbes in CI patients (P2, P3, P4, and P5) impairs wound healing time versus healthy control subjects (P1), we wounded, by 5 mm punch biopsy, CI and control patients and observed their wound healing times in the absence or presence of oral β-lactam antibiotics (carbapenems and cephalosporins) (Fig. 4D, E). In the absence of antibiotics, the time required for wound healing was significantly delayed in P2, P3, and P4 CI patients compared to healthy controls (P1), with a smaller degree of wound healing delay in P5. In contrast, administration of β-lactam antibiotics led to nearly complete wound healing in all 4 CI dysbiosis clusters (P2–P5), with little change in the trajectory of healing time by healthy controls, indicating that the altered CI microbiome impairs wound healing.
To further characterize the response of wounded skin in CI patients, we evaluated expression levels of the anti-microbial peptide β-Defensin 2 (HBD-2) in healthy subjects (P1) and CI patients (P2–P5) (Additional file 7: Fig. S6B). We observed elevations of HBD-2 mRNA in wounded skin of CI patients from dysbiosis clusters P2, P3, and P4 compared to wounded skin of healthy controls (P1). CI patients in cluster P5, however, had similar levels of HBD-2 mRNA in wounded skin as the healthy controls. S. aureus colony counts (CFU) were significantly reduced in the wounded skin of healthy subjects (P1) and P5 CI patients after approximately 36 h of healing time (Fig. 4F). S. aureus colony counts remained high in P2–P4 CI patients after 36 h. S. epidermidis colony counts among healthy subjects (P1) and CI patients (P2–P5) were similarly reduced after a wound healing time of 24–36 h (Fig. 4G), although healthy subjects started and ended with higher overall S. epidermidis colony counts. P. aeruginosa colony counts were highly increased at 2 h and reduced back at 16 h in P3 and P4 CI patient wounds, and to lesser extend in P5, from 2 to 16 h healing time (Additional file 7: Fig. S6C).
In summary, CI patients showed: (i) distinct microbiota signatures with high dysbiosis, (ii) pathogenic bacterial levels linked to IV (P2) and EI and TTD (P3) patients, and (iii) generally low levels of all microbes present in HI and LI patients (P4). Viral susceptibility, such as to HPV or HBV/HCV, is particularly linked with CI patients (Additional file 15: Table S2). Breached wounds are a key determinant of all infectious microbe levels, mainly in the ratio of S. aureus to S. epidermis (Fig. 4C and Additional file 14: Table S1) and may drive the sepsis seen in CI patients. Therapies for CI skin can correct the ratio of S. aureus to S. epidermis, as we observed for TTD patients receiving phototherapy and IV patients receiving topical ammonium bituminosulfonate (Ichthammol) 20% ointment (Additional file 9: Fig. S8).
Imbalance of immune and cytokine inflammation in CI patientsTo characterize inflammatory responses in CI patients across the dysbiosis clusters, we measured immune cell levels and cytokine responses. First, levels of granulocytes (neutrophils, eosinophils, and basophils) were measured from blood acquired from CI patients (no skin wounding; P2–P5) and healthy controls (P1). There was a statistically significant elevation in neutrophil count for all CI clusters (P2–P5) compared to healthy individuals (P1) (Fig. 5A, left panel). Interestingly, only CI patients from P4 (LI and HI) demonstrated a significant elevation in eosinophil counts (Fig. 5A, center panel), whereas there were no differences in basophils counts among CI patients compared to healthy controls (Fig. 5A, right panel).
Fig. 5
Immunotyping profile of each CI dysbiosis cluster. A Cell counts of granulocytes including neutrophils (left), eosinophils (center), and basophils (right) from healthy controls (P1) and CI patients (P2-P5) as determined by flow cytometry. Results are expressed in dot plot with each horizontal bar representing the mean of the group with statistical significance as *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001. B Cell counts and relative percentages of lymphocytes including T, B, Th17, Th1, Th2, and Treg cells as determined by flow cytometry for healthy controls (P1) and CI patients (P2–P5). Results are expressed in dot plot with each horizontal bar representing the mean of the group with statistical significance as *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001. C Correlation between SCORAD index with percentages of Th17, Th1, and Th2 cells in CI patients. Results are expressed in dot plot
Similarly, levels of T- and B- cells were quantified from blood from CI and healthy patients (Fig. 5B). Percentage of Th2+/CD4+ and Th17+/CD4+ cells are significantly increased in CI patients (P2, P3, P4, and P5) compared to healthy controls (Fig. 5B, upper right panels). Total Th1+/CD4+ cells are significantly increased in P3 CI patients, while Treg+/CD4+ cells are significantly increased in P2, P3, and P5 CI patients compared to healthy controls (P1) (Fig. 5B, bottom right panels). B cells were significantly induced in P2, P4, and P5 CI patients compared to P1 (Fig. 5B, bottom left panel). Importantly, CI patients were clinically subdivided into severe- (score > 40), moderate- (score = 15–40), and mild- (< 15) SCORAD index [27], a clinical tool for scoring the severity of atopic dermatitis/eczema; here it is applied to CI. SCORAD index in our CI cohort positively correlated with the percentage of Th17 and Th2 cells (Fig. 5C).
To further investigate our cohort for relevant genotype-immunologic phenotype correlations, Fisher exact tests and odds ratio calculations were performed for 6 immune cell types against the 7 genetic CI subtypes (Additional file 19: Table S6). We primarily observed statistically significant association of FLG mutation with neutrophil, B cell, Th1, Th2, Th17, and Treg counts; ABCA12 mutation was also associated with neutrophil elevation.
Next, we measured circulating cytokine mRNA levels of Th1 (IFN-γ, TNF-α), Th2 (IL-4, IL-5, IL-13, CCL18), Th17 (IL-1β, IL-6, IL-17A, IL-17F, IL-22, CCL20) and Treg (IL-10, TGF-β) cytokines from peripheral-blood leukocytes (Fig. 6). CI patients (P2-P5) were generally found to have higher mRNA levels of Th17 cytokines compared with healthy control (P1) (Fig. 6A), except for notable insignificance in IL-22 in P3 and IL-1β, IL-6, IL-17A, IL-17F, CCL20 in P5 compared with healthy control (P1) in one-way ANOVA analysis (Additional file 10: Fig. S9A). Similarly, the mRNA level of IL-4, IL-5, IL-13 and CCL18 of Th2 was induced in CI patients (P2, P3 and P5) (Fig. 6B), but mRNA levels of IL-13 were not significantly elevated in P5 compared to P1 nor for any of the Th2 cytokines in P4 compared to P1 (Additional file 10: Fig. S9B). In addition, an increase in the mRNA level of Th1 cytokines IFN-γ and TNF-α was detected in P3 compared to P1 (Fig. 6C), and P3 levels of IFN-γ and TNF-α were significantly higher than the other dysbiosis clusters (P1, P2, P4, and P5) (Additional file 10: Fig. S9C). The mRNA levels of IL-10 and TGF-β Treg cytokines were increased in P2, P3, and P5 compared to P1, but not P4 (Fig. 6D). These cytokine elevations were significantly highest in P3 when compared with P1, P2, and P5 (Additional file 10: Fig. S9D).
Fig. 6
Immunocytokine profiles of CI patients analyzed across the CI dysbiosis clusters. A–D The measured mRNA expression levels of Th17 (A), Th2 (B), Th1 (C), and Treg (D) cytokines to housekeeping gene (hARP) across P1-P5 dysbiosis clusters. Results are expressed in bar graphs with overlapping dots representing each individual. Bars show mean ± SEM. Kruskal Wallis tests were used to compare the mRNA expression levels between groups with statistical significance as *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001
To further analyze the signaling pathways in CI patients, we evaluated the levels of phosphorylated STAT3 Tyr705 (pY705) as a marker of Janus kinase (JAK) signaling activity in peripheral blood mononuclear cells (PBMCs) (Figs. 7 and Additional file 11: S10). Levels of STAT3 pY705 were significantly elevated in CI patients as a group compared to healthy controls (Fig. 7A). Analysis by dysbiosis cluster revealed profound elevations in STAT3 pY705 in P2, P3, and P4 compared with P1, but not in P5 (Fig. 7B).
Fig. 7
STAT3 pY705 levels as a marker for JAK/STAT signaling in CI patients. A Dot plot depicts STAT3 pY705 measured by flow cytometry in subcellular PBMCs. Line bar represents mean. Control samples were unstimulated or stimulated for 15 min with 400 μl Combo (cytokines (100 ng/ml), LPS (10 μg/ml; Sigma-Aldrich) or PMA (100 ng/ml; Sigma-Aldrich)). Healthy and CI patient samples were unstimulated by any substances. Results are plotted in dot plot showing median with interquartile range and statistical significance as *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001. B Dot plot depicts STAT3 pY705 analyzed across the 4 CI dysbiosis clusters (P2, P3, P4, and P5) compared with healthy controls (P1). Results are plotted in dot plot showing median with interquartile range and SEM statistical significance as *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001
Having established that CI patients generally display elevated neutrophils, Th17 cells, pro-inflammatory cytokines (IL-1β, TNF-α, IL-6, IL-10, and IL-17A/F), and activated JAK signaling/STAT3 p705 compared to healthy controls, we next investigated how the immune response is affected by wounding and wound healing in CI patients. Granulocyte and monocyte populations were analyzed by flow cytometry at baseline (0 h), 16 h, and 24 h after skin wounding using a 5 mm punch biopsy (Additional file 12: Fig. S11). Gr1+ granulocytes were elevated across P1-P5 at 16 h after wounding, but by 24 h post-wounding all returned closer to baseline except P2 and P3 (Additional file 12: Fig. S11B). This suggests prolonged inflammation in P2 and P3 CI patients post-wounding. Similarly, levels of Ly6+ monocytes increased across P1-P5 at 16 h post-wounding, and remained more elevated among P2, P3, and P5, and less so P4, compared to P1 at 24 h post-wounding (Additional file 12: Fig. S11C).
To further probe wound healing response in CI patients, we examined specifically the neutrophil, Th17/Treg, and dopamine-2 receptor (D2R) neural antibody counts in CI patients and healthy controls over 24 h, 48 h, and 7 days, respectively, post-wounding with a 5 mm punch biopsy (Additional file 12: Figs. S11D-F). There was a statistically significant higher proportion of neutrophils in CI patient clusters P2, P3, and P4 versus healthy controls (P1) at 24 h post-wounding, but not for CI cluster P5 (Additional file 12: Fig. S11D). The Th17/Treg count ratio was statistically elevated for all CI patients (P2-P5) compared to healthy controls at 48 h post-wounding (Additional file 12: Fig. S11E), indicating skin wounding in CI patients leads to a prolonged Th17 inflammatory response. Lastly, we examined D2R neural antibody as a measure of itch and observed statistically significant elevations in D2R levels before wounding as well as 7d post-wounding across CI patients in clusters P2-P5 (Additional file 12: Fig. S11F).
留言 (0)