
記住我




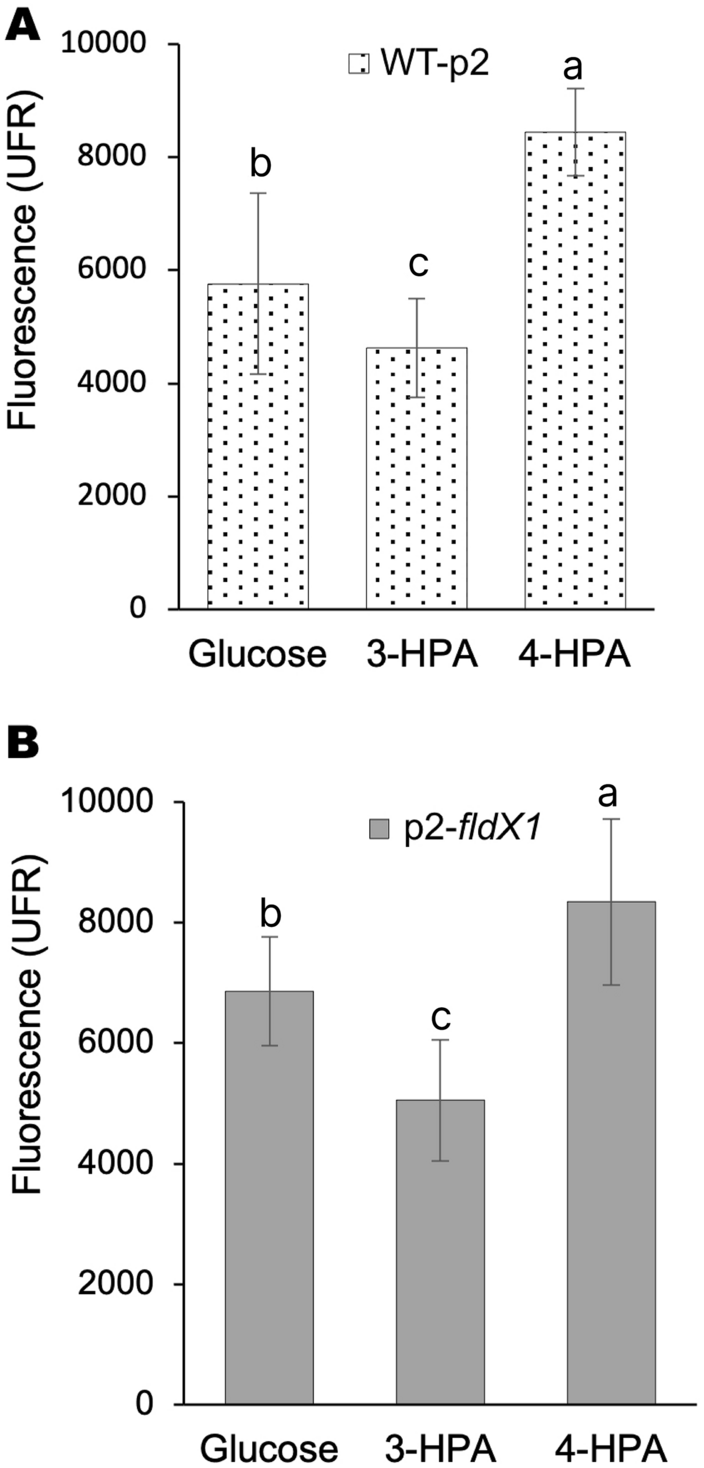

To determine how the aromatic compounds HPAs, specifically 4-HPA and 3-HPA, contribute to oxidative stress, ROS formation of P. xenovorans grown on these compounds was analyzed. Figure 1 shows that incubation of both WT-p2 and p2-fldX1 cells with 4-HPA generates more hydroxyl radicals compared to 3-HPA and to glucose (control).
Fig. 1
ROS formation in P. xenovorans recombinant strains upon 4-HPA and 3-HPA exposure. Cells were grown on glucose until exponential phase and incubated with 3-HPA or 4-HPA (5 mM). A, strain WT-p2. B, strain p2-fldX1. Unexposed cells were also measured (control). Each value is an average ± SD of at least three independent experiments. Significant differences between groups were evaluated using one-way ANOVA followed by a Fischer's least significant difference (LSD) test (α = 0.05)
The flavodoxin FldX1 improves growth and degradation of 3-HPA and 4-HPATo evaluate the effect of FldX1 on the growth of P. xenovorans on aromatic compounds, the recombinant strains were grown in M9 minimal medium using 3-HPA or 4-HPA (5 mM) as sole carbon and energy sources. During growth on 3-HPA (Fig. 2A) strain overexpressing the flavodoxin FldX1 showed a higher growth rate (0.074 h−1) than the control strain (0.058 h−1). Strain p2-fldX1 reached the stationary phase after 18 h incubation, while strain WT-p2 reached this phase after 21 h. Similarly, during growth on 4-HPA (Fig. 2B), strain p2-fldX1 exhibited a higher growth rate (0.119 h−1) than strain WT-p2 (0.081 h−1). Strain overexpressing the flavodoxin FldX1 reached the stationary phase after 12 h incubation, while the strain WT-p2 reached this phase at ~ 14 h. Interestingly, both strains (p2-fldX1 and WT-p2) grew faster on 4-HPA as sole carbon and energy sources compared to 3-HPA. This result is in accordance with a previous study carried out with the wild-type strain LB400, where the culture reached a higher cell density growing on 4-HPA as sole carbon an energy source in comparison to 3-HPA [25].
Fig. 2
Effects of FldX1 on growth and degradation of 3-HPA and 4-HPA. Cells (p2-fldX1 and WT-p2) were grown for 24 h in M9 minimal medium on (A) 3-HPA or (B) 4-HPA as sole carbon and energy sources. Growth was determined measuring turbidity at 600 nm. Different letters within a time point indicate significant differences from each other according to Student’s t-test, at p ≤ 0.05. 3-HPA and 4-HPA concentrations were determined in the supernatants by HPLC–DAD. (C) 3-HPA. D 4-HPA. Different letters within a time point indicate significant differences from each other according to one-way ANOVA followed by Tuckey´s HSD
To evaluate the effect of FldX1 on the degradation capabilities of p2-fldX1 and WT-p2 strains, 3-HPA or 4-HPA were quantified in culture supernatants by HPLC–DAD. Strain p2-fldX1, overexpressing the flavodoxin FldX1, showed an improved 3-HPA degradation compared to the control strain (Fig. 2C). 3-HPA was completely degraded by strain p2-fldX1 at 18 h of incubation, whereas the strain WT-p2 completely degraded 3-HPA at 21 h. Figure 2D shows that p2-fldX1 cells also exhibited a higher 4-HPA degradation compared to the control strain. Interestingly, 4-HPA was completely degraded by the strain p2-fldX1 earlier than 3-HPA (~ 12 h), while the control strain (WT-p2) completely degraded 4-HPA at ~ 15 h. In accordance with the growth curves, both strains p2-fldX1 and WT-p2, degraded 4-HPA faster than 3-HPA. These results indicate that the flavodoxin FldX1 confers an advantage during bacterial growth and degradation of 3-HPA and 4-HPA compounds.
FldX1 prevented the upregulation of stress-related proteinsTo understand the effects of FldX1 overexpression on P. xenovorans proteome grown on 3-HPA or 4-HPA, protein expression analyses were performed using glucose as control. In this assay, a total of 3,179 proteins from P. xenovorans were quantified from a total of 8,754 coding sequences of the genome. Nevertheless, only proteins identified in ≥ 4 of the 6 replicates of each condition were considered for the statistical analysis, reducing the identified proteins to 2,093 proteins that fit our inclusion criteria (Additional file 1: Table S2 and Figure S1). Additional file 1: Table S2 contains all the relevant information of the identified and quantified proteins from the mass spectrometry analysis. Besides the functional annotation and classification into clusters of orthologous groups (COG) of the proteins, the quantitative data can be assessed: Label-free quantification data (LFQ) obtained from the MaxQuant software indicate the abundance of the respective protein–this can be used to rank proteins according to their abundance in a strain/condition; averaged values of LFQ and the calculated fold changes can also be drawn from this table. Also, the result of the statistical analysis obtained by the MaxQuant companion software Perseus (t-test) is indicated for each protein. The Additional file 1: Fig. S1 visualize an upset plot showing the shared and unique proteins in the different samples.
The log2-fold change indicates the change in comparison to the glucose-grown strain. The (log2) fold change is calculated based on average LFQ values within the replicates for each strain/condition. If these quantitative values within the replicates of a strain/condition vary much, this impairs statistical significance. As a result, the difference (fold change) is not significant although the averaged values lead to high induction factors. Additional file 1: Table S2 contains a column with bar charts indicating the LFQ values of each sample and allowing easy comparison of the variances of these LFQ values.
The analysis focused on comparing each strain (p2-fldX1 and control) grown on 3-HPA or 4-HPA versus glucose-grown cells (control). A very low induction/repression ratio might not be biologically relevant even if it is statistically significant. Proteins with significantly changed amount and a log2-fold change of 1 (or − 1 for repression; corresponding to twofold change) are highlighted in the Additional file 1: Table S2 for better visualization to account for biological relevance. Proteome analysis revealed novel information about the antioxidant stress response and the physiological response to HPA catabolism. In p2-fldX1 cells cultured in 3-HPA, 109 and 83 proteins were upregulated and downregulated compared to glucose-grown cells, respectively. When p2-fldX1 cells were cultured on 4-HPA, 128 and 85 proteins were upregulated and downregulated, respectively. Of all these proteins, 24 and 26 of them were only detected in p2-fldX1 cells during growth on 3-HPA and 4-HPA (“on”), respectively, but not in the control condition (Additional file 1: Table S3). Similarly, 21 and 25 proteins were not detected in p2-fldX1 cells when cultured on 3-HPA and 4-HPA, respectively, in comparison to glucose (“off”) (Additional file 1: Table S4).
In WT-p2 cells growing on 3-HPA, 104 and 80 proteins were upregulated and downregulated, respectively, compared to glucose-grown cells. When strain WT-p2 was cultured on 4-HPA, 171 and 125 proteins were upregulated and downregulated, respectively. Of all these proteins, 24 and 24 were “on” during growth on 3-HPA and 4-HPA, respectively, and 21 and 24 were “off” during the same conditions, compared to glucose.
Upregulation of the 3-HPA and 4-HPA catabolic proteinsOf the proteins induced in p2-fldX1 cells during HPA growth condition, aromatic catabolic proteins, and transport proteins were identified. Several enzymes of 3-HPA and 4-HPA degradation pathways were identified by proteomics. Upregulation of the 5-carboxymethyl-2-hydroxy-muconic semialdehyde (CHMS) dehydrogenase (HpaE, Bxe_B2030) protein from the homoprotocatechuate (HPC) pathway was detected on strains p2-fldX1 and WT-p2 during growth on 3-HPA and 4-HPA (Fig. 3). HpaE transforms CHMS in 5-carboxymethyl-2-hydroxy-muconic acid (CHM). The HpaD protein (Bxe_B2031), the ring-cleavage dioxygenase of the homoprotocatechuate pathway, was not detected in the proteomic analysis. Other enzymes of this pathway were not identified or did not fit the filter criteria for quantification. In the case of the homogentisate (HMG) pathway, the subunit A (MhaA, Bxe_A2727) of the enzyme 3-HPA 6-hydroxylase (MhaAB) was upregulated in both strains during growth on 3-HPA and 4-HPA, although no significant statistically difference was calculated. Interestingly, in the case of strain WT-p2 cultured in 4-HPA, the expression of the protein was very low compared to 3-HPA. The subunit B of this (MhaB, Bxe_A2726) was not detected. The same effect was identified for the first copy of the enzyme HMG 1,2-dioxygenase (HmgA1, Bxe_A2725). The first copy of the maleylacetoacetate isomerase (HmgC2, Bxe_A4141) was significantly upregulated in both strains p2-fldX1 and WT-p2 cultured in 4-HPA. Nevertheless, the other copy of this enzyme (HmgC1, Bxe_A2723) did not show changes in expression levels (Fig. 3). The two copies of the enzyme fumarylacetoacetate hydrolase (HmgB1 (Bxe_A2724) and HmgB2 (Bxe_A3899)) were upregulated in both strains growing on 3-HPA and 4-HPA. The induction of Hmg proteins in presence of 3-HPA and 4-HPA indicates the utilization of the homogentisate ring-cleavage pathway for the degradation of both HPAs.
Fig. 3
Overview of the protein expression profile for the catabolism of 3-HPA and 4-HPA in P. xenovorans strains. Heatmaps show changes in protein expression levels of cells grown until exponential phase on 3-HPA and 4-HPA relative to glucose as sole carbon source. Each value is an average of Log2(expression change) of at least four independent experiments. Significant statistically differences are represented as colored border for the corresponding strain. Peripheral and central pathways for 3-HPA and 4-HPA are depicted including their metabolic intermediates. Catabolic enzymes: MhaAB: 3-HPA 6-hydroxylase; HmgA: HMG 1,2-dioxygenase; HmgB: Fumarylacetate hydrolase; HmgC: Maleylacetate isomerase; HpaBC: 4-HPA monooxygenase; HpaD: HPC 2,3-dioxygenase; HpaE: CHMS dehydrogenase; HpaF: CHM isomerase; HpaG: OPET decarboxylase. Metabolic intermediates of 3-HPA and 4-HPA degradation: HMG: Homogentisate.; MA: Maleylacetoacetate; FA: Fumarylacetoacetate; HPC: Homoprotocatechuate; CHMS: 5-carboxymethyl-2-hydroxy-muconic semialdehyde; CHM: 5-carboxymethyl-2-hydroxy-muconic acid.; OPET: 5-oxo-pent-3-ene-1,2,5-tricarboxylic acid; HHDD: 2-hydroxy-hept-2,4-diene-1,7-dioic acid. TCA cycle: Tricarboxylic acid cycle. ND: Not determined
Interestingly, during the growth of strains p2-fldX1 and WT-p2 on 3-HPA and 4-HPA, almost all proteins of the phenylacetic acid degradation pathways described in P. xenovorans LB400 [38] were upregulated, with higher expression levels observed in 3-HPA. Catabolic proteins of the phenylacetate pathway (paaH, paaI and paaJ) were induced in presence of 3-HPA and 4-HPA. In addition, transcriptional analyses during growth on 3-HPA and 4-HPA, showed that during growth on 3-HPA (Additional file 1: Fig. S2), p2-fldX1 and WT-p2 exhibited an expected increase of hmgA1 gene transcript compared to growth on glucose (199- and 328-fold, respectively). In 3-HPA-grown cells, hmgA2 gene increased 121- and 176-fold in the strains p2-fldX1 and WT-p2, respectively. Interestingly, for both genes hmgA1 and hmgA2, strain WT-p2 exhibited higher expression levels than strain p2-fldX1. This suggests that downregulation of catabolic genes in strain p2-fldX1 may be a consequence of a minor oxidative stress condition and an improved catabolic performance, causing a decrease of intracellular 3-HPA concentration. The hpaD gene in both strains p2-fldX1 and WT-p2 did not show variations in expression levels in comparison to the control, and showed low expression levels (Additional file 1: Fig. S2). This is in accordance with proteomic analyses, in which undetectable HpaD protein levels were observed probably below detection limit for MS analysis. 4-HPA catabolism showed a similar expression increase of the hmgA1 gene expression for both strains (128 to 130-fold), compared to glucose-grown cells (Additional file 1: Fig. S2). No changes were detected in hmgA1 and hpaD gene expression levels, suggesting that the higher degradation by strain p2-fldX1 is not based on the differential expression of these ring-cleavage enzymes of the catabolic pathway.
Other proteins that probably are related to aromatic catabolism were induced in both strains (p2-fldX1 and WT-p2) in 3-HPA and 4-HPA: an extradiol ring-cleavage dioxygenase (Bxe_A0432), an aldehyde dehydrogenase (Bxe_B2718) and an alkanesulfonate monooxygenase (Bxe_A2493) (Additional file 1: Table S3). A FAD-dependent oxidoreductase (Bxe_B1666) was upregulated in p2-fldX1 in 3-HPA; a salicylate dehydrogenase (Bxe_C0213) was induced in p2-fldX1 in 4-HPA and in WT-p2 in both aromatics; and a shikimate 5-dehydrogenase (Bxe_B0884) was upregulated in both strains in 4-HPA.
Upregulation of antioxidant proteinsFigure 4 shows changes of antioxidant-related proteins during growth of p2-fldX1 and WT-p2 strains in 3-HPA or 4-HPA, compared to growth on glucose. In general, the catabolism of 4-HPA produced more protein changes than 3-HPA compared to glucose grown-cells (Fig. 4), suggesting that growth on 4-HPA may generate an increased oxidative stress response. This tendency is more significant when the proteins with a significant change against control were evaluated. ROS scavenger enzymes AhpC2, AhpF, AhpD3, KatG, KatA, CpoF and OsmC were upregulated in both conditions (3-HPA and 4-HPA), however, higher expression levels were observed in 4-HPA-grown cells. Similarly, an increased downregulation of DpsA, a protein from the ferritin family that protects against oxidative stress, was observed in 4-HPA-grown cells. A significant difference is only observed in 4-HPA-grown cells for the alkyl hydroperoxide reductase system subunit AhpD3, and the organic hydroperoxide resistance protein OhrB, where no significant differences were observed in 3-HPA-grown cells compared to the glucose-grown cells. Overall, a slight repression of stress-related proteins was observed in p2-fldX1 cells compared to control cells (WT-p2), suggesting that overexpression of FldX1 prevents the antioxidant response due to a protective effect.
Fig. 4
Changes in antioxidant proteins levels of P. xenovorans during growth on 3-HPA and 4-HPA. Heatmaps show changes in protein expression levels of cells grown until exponential phase on 3-HPA or 4-HPA compared to glucose-grown cells. Only proteins with variation in the expression levels are represented. Each value represents the average of Log2(expression change) of at least four independent experiments. Significant differences are represented as colored border for the corresponding strain. TR: Transcriptional regulator. OS: Oxidative stress. GstA or GstB: Glutathione S-transferase. Grx: Glutaredoxin. GorA: Glutathione peroxidase. ArsC2: Arsenate reductase (glutaredoxin). BphK: Glutathione S-transferase (biphenyl degradation pathway). YgjG: Glutathionyl-hydroquinone reductase. TrxA: Thioredoxin. TrxB: Thioredoxin reductase. FumC: Fumarate hydratase class II. AcnA: Aconitate hydratase. AhpC or AhpD: Alkyl hydroperoxide reductase, peroxidase subunit. AhpF: Alkyl hydroperoxide reductase, reductase subunit. Kat: Catalase. SodB: Superoxide dismutase. Prx or Bcp: Peroxiredoxin. YfeX: Dye-decolorizing peroxidase. CpoF: Non-heme chloroperoxidase. Tpx: Thiol peroxidase. YghU: Organic hydroperoxidase. OsmC: Peroxiredoxin. FldX1: Flavodoxin. Fpr: Ferredoxin NADP reductase. RubA1: Rubredoxin. DpsA: Ferritin DPS family DNA-binding protein. Hpf: High potential Fe-S protein. EfeN: Deferrochelatase/peroxidase. OhrA or OhrB: Organic hydroperoxide resistance protein
A similar protein expression profile related to oxidative response was observed when strains p2-fldX1 and WT-p2 were compared on the same condition (3-HPA or 4-HPA). For 4-HPA, two thiol-redox homeostasis proteins GstB3 and TrxA3, and a ROS-resistant isoenzyme (AcnA2) were upregulated in strain WT-p2 in comparison to glucose (Fig. 4). Peroxiredoxins Prx1, Prx2, Bcp, and the high-potential iron-sulfur protein Hpf were downregulated in WT-p2. In 3-HPA-grown WT-p2 cells, only two proteins showed a significant difference in expression levels: the peroxiredoxin OsmC was upregulated and the organic hydroperoxide resistance protein A (OhrA) was downregulated in strain WT-p2 grown on 3-HPA. These results suggest that overexpression of the flavodoxin FldX1 leads to a differential response to oxidative stress in both aromatic compounds, observing a higher stress response towards oxidative damage on strain WT-p2 during the growth on 4-HPA.
Several other proteins displayed expression changes. General stress proteins chaperonin GroEL (Bxe_B1569) and a cold-shock protein (Bxe_B2780) were induced during growth of strain p2-fldX1 on 3-HPA, but not on 4-HPA.The 50S ribosomal protein L36 (Bxe_A0336) were downregulated in the control strain cultured in 3-HPA.
More than 20 membrane proteins, including porins and transporters, showed changes in expression during growth of the strains p2-fldX1 and WT-p2 in the aromatic compounds. Some of these proteins were induced or repressed in different conditions. Proteins FliC, FlhF, FlgL and FliJ (Bxe_A0103, Bxe_A0132, Bxe_A0151 and Bxe_A0162, respectively), involved in flagella biosynthesis and functionality, were downregulated or “off” in the recombinant strain growing on 3-HPA and/or 4-HPA (Additional file 1: Table S4). Fimbria/pilus periplasmic chaperone (Bxe_B2971) was “on” in strains p2-fldX1 and WT-p2 cultured on 3-HPA and 4-HPA. Two major facilitator superfamily (MFS) transporters (Bxe_A3901 and Bxe_B0430), a dicarboxylate periplasmic transporter (Bxe_B0438; DctP family), a sugar ABC transporter (Bxe_A2975), and an OmpC family outer membrane porin were induced only in presence of 3-HPA and 4-HPA, but not on glucose-grown cells (“on”) (Additional file 1: Table S3).
FldX1 decreases gene expression of oxidative stress-related transcripts during growth on HPAsPreviously identified proteins associated with oxidative stress mechanisms in strain LB400 [4] were evaluated in detail to compare the oxidative stress response of both strains. To evaluate if the oxidative stress mechanisms were differentially induced in the recombinant strains, gene expression of transcripts involved in the antioxidant response were analyzed by qRT-PCR during growth on 3-HPA and 4-HPA. The genes selected included some of the ROS scavenging enzymes previously identified as part of the antioxidant machinery of strain LB400, such as the alkyl hydroperoxide reductase (ahpC1), catalase (katE) and superoxide dismutase (sodB1), along with the OxyR transcriptional regulator (oxyR), two thioredoxin reductases (trxB1 and trxB2), an organic hydroperoxide resistance protein (ohrB), a glutathione S-transferase (gstA1), a high potential iron-sulfur protein (electron transfer protein; hpf) and the ROS resistant isoform of the fumarate hydratase (fumC) [4, 29]. No > twofold changes in gene expression levels were observed in cells gown on 3-HPA and 4-HPA compared to glucose (Fig. 5). Nevertheless, strain p2-fldX1 showed a slight tendency to decrease the expression of specific genes (i.e., katE and fumC in 3-HPA-grown cells; katE, sodB1, trxB1 and ohrB in 4-HPA-grown cells). Interestingly, in both growth conditions, the transcript levels of these genes were higher in the strain WT-p2, whereas lower expression levels were observed in strain p2-fldX1, suggesting that FldX1 overexpression might exert an adjuvant role, wherein the expression of these stress-related genes is nonessential, or contributing to a more favorable redox environment so their expression is not required.
Fig. 5
Effects of flavodoxin FldX1 on the expression of antioxidant response genes in P. xenovorans during growth on 3-HPA and 4-HPA. Bar plots show changes in expression levels of cells grown until exponential phase on A, 3-HPA and B, 4-HPA relative to glucose-grown cells. Genes ftsZ and gyrB were used as reference genes. Each value is an average ± SD of at least three independent experiments
FldX1 increases early 4-HPA degradation in soil microcosms assaysTo evaluate potential use of the strain overexpressing the flavodoxin FldX1 in bioremediation processes, soil microcosms assays polluted with 4-HPA were carried out. 4-HPA was selected for this study due to the overall catabolic fitness of the strain p2-fldX1 on this aromatic compound. Moreover, higher ROS levels were observed upon exposure to 4-HPA (Fig. 1).
An increase in 4-HPA degraders was observed within 14 days of incubation, especially under conditions bioaugmented with strain p2-fldX1 in the presence of 4-HPA (Fig. 6A, B). All bioaugmented treatments showed a complete 4-HPA degradation at the end of the experiment (Fig. 6C). Notably, strain overexpressing the flavodoxin FldX1 degraded more 4-HPA than the control strain at 3 and 7 days of incubation. In accordance with these results, in presence of 4-HPA (day 21), higher levels of 4-HPA degrading bacteria were observed in soils bioaugmented with strain p2-fldX1 compared to soils inoculated with WT-p2 (Fig. 6A). In the uninoculated control, colonies were observed from day 14 until the end of the experiment (21 days). Total heterotrophs increased CFU counts per g of dry soil after 14 d incubation, especially in bioaugmented soils (p2-fldX1 and WT-p2) spiked with 4-HPA (Additional file 1: Fig. S3).
Fig. 6
Effect of flavodoxin FldX1 in P. xenovorans on 4-HPA degradation in soil microcosms. A 4-HPA degraders and (B) 4-HPA degraders resistant to kanamycin. (C) 4-HPA degradation in soil microcosms. D Prevalence of plasmids from P. xenovorans recombinant strains in soil microcosms. Significant differences were determined using Fisher's LSD test (p < 0.05). Lowercase letters above the error bars indicate significant differences between treatments. Asterisks indicate value = 0. Each value is an average ± SD of the three replicates of the experiment
As strains p2-fldX1 and WT-p2 possess a plasmid with kanamycin resistance as a selection marker, the plasmid was monitored during incubation using M9 minimal medium with the antibiotic and 4-HPA as sole carbon and energy sources (Fig. 6B, D). No growth was detected in the uninoculated condition. In presence of the aromatic compound, the bioaugmented condition with the strain overexpressing the flavodoxin FldX1 exhibited higher growth than the control strain. Interestingly, the recombinant strains p2-fldX1 and WT-p2 did not show growth differences between them in the absence of 4-HPA. Regarding the plasmid loss, indicated by the decrease in kanamycin resistance, this phenomenon is evident in all the bioaugmented treatments. In presence of 4-HPA, the strain overexpressing the flavodoxin FldX1 showed a lower plasmid loss at days 7 and 21, compared to the control strain. In the absence of the aromatic compound, strain p2-fldX1 exhibited a higher quantity than strain WT-p2 at day 21. At the end of the assay, only 10 to 30% of plasmid remains (Fig. 6D).
留言 (0)