Early embryonic development thrives within the optimal microenvironment provided by the oviduct. In cattle, the embryo spends four days in contact with oviductal epithelial cells and the various components of the oviductal fluid, including EVs. These vesicles are potentially involved in maternal communication with gametes and embryos (reviewed in Cajas et al. [1], as they transfer biomolecules such as miRNAs that can modulate the activities of recipient cells.
Previous studies have demonstrated that in vivo-derived EVs from the bovine oviduct can traverse the zona pellucida (ZP) and be taken up into the cytoplasm of blastocyst cells. This suggests that molecular cargos in oviductal EVs can be incorporated into the developing embryo, as observed in studies by Almiñana et al. [7] and Leal et al. [25]. Furthermore, it has been shown that the addition of oviductal EVs to in vitro culture media enhances embryo quality and blastocyst hatching rate in bovine embryos [7].
Along the same lines, we demonstrated that EVs from bovine oviductal epithelial cell (BOEC) conditioned media can be isolated, characterized, and successfully used for in vitro embryo culture, resulting in improved blastocyst quality [43]. Additionally, supplementation of in vitro culture media with EVs from oviductal fluid during in vitro culture enhances development and increases blastocyst quality in terms of cryotolerance and the expression patterns of quality-related genes [6].
Bauersachs et al. [44] showed that supplementation of in vitro culture media with EVs from oviductal fluid alters the embryonic transcriptome of bovine blastocysts, leading to a decrease in apoptosis of embryonic cells and improved embryo viability. Furthermore, specific miRNAs identified in oviductal EVs can modulate the expression of certain genes. For instance, Sp3 Transcription Factor expression can be targeted by miR-27a-3p, miR-484, miR-1260b, and miR-218-5p; and NANOG can be targeted by miR-34a-5p, miR-34c-5p, miR-34b-3p, miR-335-5p, miR-128-3p, miR-150-5p, and miR-125b-2-3p [44].
Recently, our group demonstrated that mimicking physiological conditions using EVs from OF and UF in sequential IVC does not affect embryo development but significantly improves blastocyst quality. The observed improvements include enhanced survival rates after vitrification/warming, increased total cell number, altered lipid content, and relative changes in the expression of lipid metabolism transcripts and lipase activation. The contents of EVs, particularly miRNAs, may contribute to these observed effects. Notably, our results highlights the increased abundance of microRNA 148b (bta-miR-148b) in EVs derived from oviductal fluid collected during the early luteal phase (Day 1 to 4) in heifers [25]. MiR-148b-3p plays a significant role in cell proliferation, migration, and invasion in human cancer cells, and its effect depends on the level of expression [26, 27]. Moreover, the downregulation of miR-148b-3p in both human and mouse frozen-thawed sperm affects its expression during subsequent embryonic development, resulting in a significantly lower embryo formation rate [45].
Here, we verified that synthetic miR-148b (designed to replicate naturally occurring, mature miRNA), when supplemented to the culture medium, is taken up by embryos of < 16 cells and BD7 through passive transfection. We also validated SMAD5, an essential transcriptional effector of the BMP/TGF-β pathway involved in embryonic development, as a potential target gene for miR-148b. In our functional study, we observed that the presence of miR-148b during the entire culture period (Day 1 to Day 7) or from Day 1 to Day 4 (representing the effect of miR-148b when the embryo is in the oviduct) resulted in an improvement of embryo quality. While embryo development remained unaffected, we noticed an increase in the total number of cells, trophoblast (TE) and inner cell mass (ICM) cells, along with a higher number of cells labeled with POUF51. Moreover, miR-148b exhibited a dual effect by modulating the relative abundance of key genes associated with cellular differentiation and proliferation in the BMP/TGF-β signaling pathway, while also increasing the level of p-SMAD1/5, thus confirming its influence on this pathway. This study suggests that the addition of miR-148b to in vitro culture media enhances embryo quality, indicating its potential significance as a key signaling molecule for embryo-maternal interaction in the oviduct during the preimplantation period. Furthermore, it highlights the involvement of extracellular vesicles in delivering miR-148b.
Besides, our data revealed that fluorescently tagged miRNA mimics can be incorporated by embryos without the addition of transfection reagents. This phenomenon, known as “gymnosis” or passive transfection, is a poorly understood process through which various cell types can internalize miRNAs in vitro without association with protein complexes or the addition of transfection reagents [46, 47]. Previous studies have also reported this uptake mechanism in bovine embryos [5]. Furthermore, we confirmed through qPCR that the miR-148b mimic is indeed taken up by 16-cell embryos and blastocysts, as evidenced by significantly higher levels (approximately 50 times higher) compared to control groups. These findings align with a study by Lin et al. [23], which demonstrated the uptake of miR-10b by embryos, where qPCR results exhibited notably higher levels (approximately 70 times higher) compared to the control group.
Based on computational algorithm analysis, we evaluate the modulation of putative target genes of miR-148b in cattle. The results revealed that the mRNA abundance of SMAD5 decreased in 16-cell embryos and Day 7 blastocysts from the miR-148b group compared to the control group. Hence, our study suggests that miR-148b directly targets SMAD5 and inhibits the transcription of this gene. MiRNAs predominantly function to reduce target mRNA levels [48]. This reduction in mRNA levels is associated with poly(A) tail shortening, suggesting a model in which miRNAs induce mRNA de-adenylation. This de-adenylation process promotes decapping and facilitates more rapid degradation through standard mRNA turnover processes [49,50,51].
To further comprehend the mechanisms by which miR-148b affects early embryonic development, we designed a functionality test by adding a specific miRNA mimetic (miR-148b) to the embryo culture medium. Our results are consistent with previous studies that have demonstrated that the addition of miRNA mimics to the embryo culture medium does not adversely affect development [5, 23]. In contrast, a recent study by Aoki et al. [52] reported enhanced embryonic development following the supplementation of miRNA mimics in the culture media. This suggests that the efficacy of improving in vitro embryo production might be contingent upon the specific functional role exerted by the added miRNA.
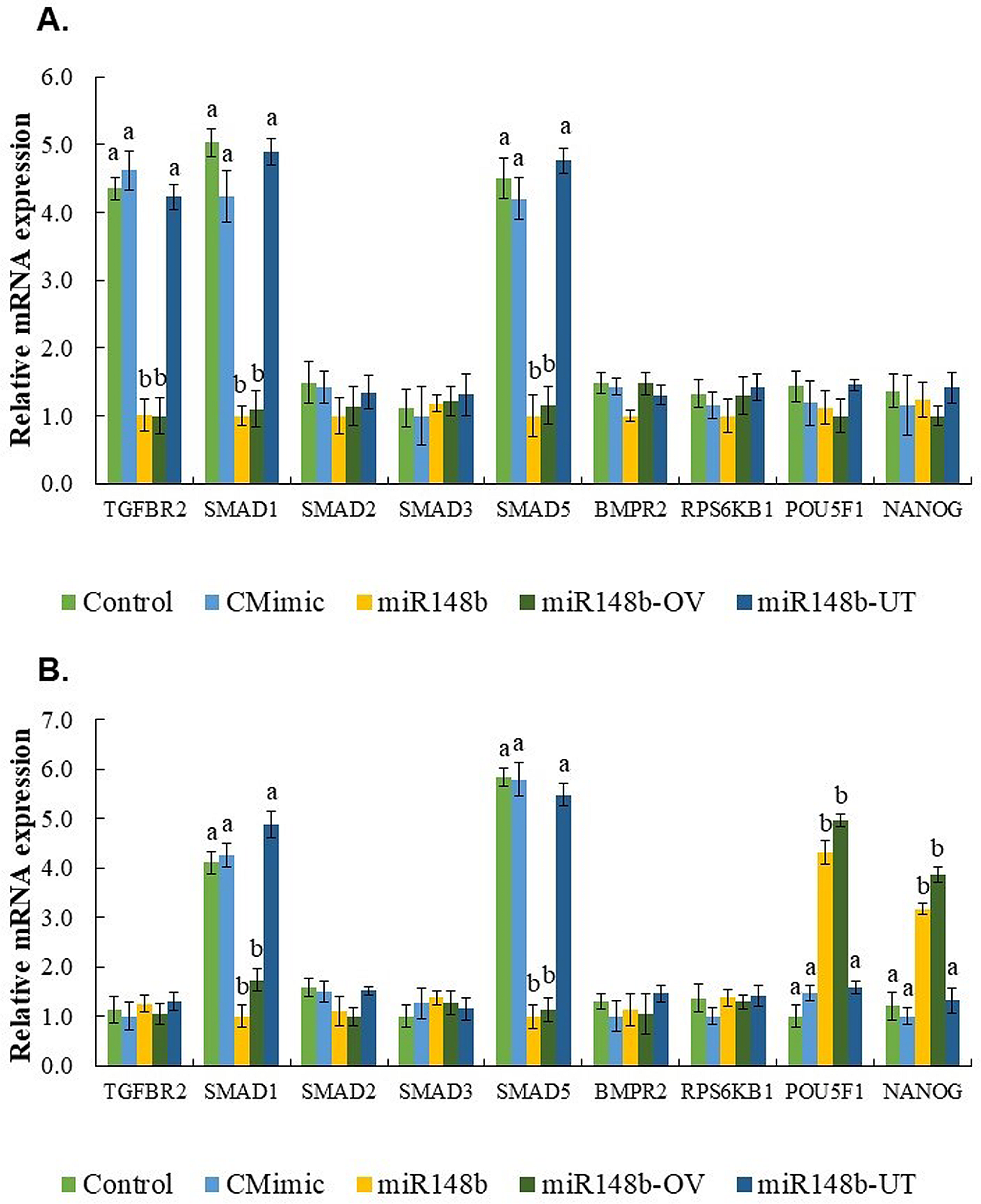
On the contrary, miR-148b exhibited a notable improvement in various embryo quality parameters within the miR-148 (supplemented throughout the entire period of IVC) and miR148-OV (supplemented during days 1 to 4 of IVC, representing the miRNA’s effect in the oviduct) groups. Specifically, it led to a significant increase in the total number of cells, trophoblast (TE), inner cell mass (ICM) cells, and POUF51-labeled cells. Furthermore, miR-148b demonstrated its regulatory role by influencing the expression of key genes involved in cellular proliferation and differentiation, such as POU5F1 and NANOG.
During mammalian preimplantation development, OCT4/POU5F1 plays a critical role as a transcription factor, with diverse functions [53], encompassing the maintenance of pluripotency and the regulation of differentiation events [54]. Our results showed that the pluripotency factor OCT4/POU5F1 is consistently expressed across the entire bovine Day 7 blastocysts, similar to the observations by Simmet et al. [55, 56] and Kirchhof et al. [57] in both in vivo and in vitro produced bovine embryos on days 7, 9, and 12. Furthermore, we observed an increased expression of POU5F1 and NANOG transcripts in blastocysts produced with miR-148b supplemented throughout the entire period of IVC or during days 1 to 4 of IVC to simulate the miRNA effect in the oviduct. Previous studies have indicated that NANOG activation does not rely on the embryonic activation of OCT4 [53, 58]. However, the absence or very low levels of the epiblast marker NANOG in OCT4 knockout (KO) blastocysts on day 7 suggests that the maintenance of epiblast cells during the early stages of the second lineage differentiation fails in the absence of OCT4, thereby confirming the requirement of OCT4/POU5F1 for NANOG expression in bovine blastocysts [55]. In another study by the same group, OCT4 KO blastocysts generated through somatic cell nuclear transfer and zygote injection demonstrated that both epiblast maintenance and hypoblast differentiation rely on OCT4. Consequently, it was concluded that OCT4 is necessary for the maintenance of pluripotency in the epiblast and the differentiation of the hypoblast during the second lineage differentiation in bovine preimplantation embryos [55, 56].
In addition, the pluripotent state in murine and human embryonic stem cells is maintained through the coordinated activity of BMP signaling, along with the transcription factors OCT4 and NANOG [59,60,61]. BMPs belong to the TGF-β superfamily and employ downstream effectors—SMAD1, SMAD5, and SMAD8—to transmit signaling. Notably, a fascinating finding suggests that NANOG physically interacts with SMAD1, thereby interfering with the recruitment of coactivators to active SMAD1 complexes and influencing the activity of BMP signaling [62]. Studies provide evidence that bovine embryos can exhibit a response to BMP2 stimulation within the initial 3 days of in vitro culture. This response manifests as an increase in blastocyst mRNA for NANOG and CDX2, which is measured 4 days later. Furthermore, the effects of BMP2 treatment on cell allocation indices are observed even after the administration of the treatment. The precise mechanisms underlying the heightened expression of blastocyst NANOG and CDX2 mRNA in response to BMP2 stimulation remain unknown. However, previous research has demonstrated that BMP4 can induce CDX2 mRNA expression [63] and promoter activity [64] in other cell lines and human ES cells. Notably, both BMP2 and BMP4 signaling pathways operate through SMAD1/5 [65]. Collectively, these results reveal the critical role of BMPs in preimplantation embryo development, which opens the possibility of establishing BMP/TGF-β as an important pathway of action for miR-148b in the bovine embryo.
The activity of TGF-β ligands is mediated predominantly through SMAD signaling [66]. Briefly, upon binding between the TGF-β ligand and type II receptor, the type I receptor is recruited and phosphorylated, which in turn activates SMADs (R-SMADs) by phosphorylation. SMAD1/5/8 are activated by BMPs, while SMAD2/3 are activated by activin and TGF-β binding to cognate receptors. Once activated, R-SMADs bind to SMAD4 and are then imported to the nucleus, where they regulate gene transcription [67, 68].
To verify the action of miR-148b on the quality of 16-cell and blastocyst stage embryos, and its possible implication in signaling the BMP/TGF-β pathway, we analyzed the expression of candidate genes like TGFBR2, a key regulator of the TGF-β pathway, and SMAD1, SMAD2, SMAD3, SMAD5, BMPR2, RPS6KB1. The results showed that the relative abundance of TGFBR2 was only decreased in 16-cell embryos for the miR148 (supplemented throughout the entire period of IVC) and miR148-OV (supplemented during days 1 to 4 of IVC, representing the miRNA effect in the oviduct) groups. Barrera et al. [69] reported that TGFBR2 expression is present from the 2- to 8-cell stage embryos but was not detected at the blastocyst stage. Unfortunately, they did not analyze the embryos at > 16-cell stage, so we speculate that the expression of TGFBR2 tends to decrease during the major phase of embryonic genome activation (MJEGA-8- to 16-cell stage); however, this needs to be verified.
Our findings revealed significant decreases in mRNA transcripts of SMAD1 and SMAD5 in 16-cell embryos and blastocysts from the miR148 (supplemented throughout the entire period of IVC) and miR148-OV (supplemented during days 1 to 4 of IVC, representing the miRNA effect in the oviduct) groups. These results are consistent with a previous study by Lee et al. [67] which reported that SMAD1 mRNA levels remained elevated after fertilization until the 4-cell stage but showed a subsequent decrease in 16-cell, morula, and blastocyst stage embryos. In contrast, SMAD5 mRNA levels increased after fertilization at the pronuclear, 2-cell, and 4-cell stages, reaching their lowest levels at the 16-cell, morula, and blastocyst stages [67]. These findings suggest that SMAD1 and SMAD5 transcripts are of maternal origin, and their downregulation may play a crucial role following major embryonic genome activation.
Moreover, miRNAs are strong candidates for facilitating maternal mRNA degradation during the maternal-to-embryonic transition (MET) due to their ability to regulate gene expression in a temporally and spatially specific manner. Several studies have demonstrated the involvement of miRNAs in the development of various species, particularly in the degradation of maternal transcripts. These studies have identified the presence of miRNAs at distinct developmental stages prior to EGA, and in all cases, miRNA profiles were found to be up-regulated during EGA [35, 50, 70]. These characteristic expression profiles strongly support the hypothesis that miR-148b undergoes processing during EGA to serve as a guide for the degradation of maternal mRNA.
In a previous study [33], we observed the dynamic regulation of ligands, receptors, and signaling components of the BMP signaling pathway during early embryogenesis in bovine. However, the presence of these transcripts does not mean that BMP signaling is active during early embryonic development. To corroborate this, we conducted immunofluorescence analysis to investigate the localization of the pSMAD1/5 protein and performed Western blot analysis to assess its expression in blastocysts. Our findings revealed that pSMAD1/5 protein, produced by BMP-activated receptor kinase, was predominantly localized in the nucleus of Day 7 blastocysts across all groups. This finding is consistent with similar observations made by Rajput et al. [71], suggesting that the activation of SMAD-dependent BMP signaling may occur during embryonic genome activation. This activation could potentially play a role in crucial developmental events such as blastocyst formation and the determination of the first cell lineage in cattle.
Our results indicated that the downregulation of SMAD5 and SMAD1 mRNA could compromise the protein levels. Surprisingly, Western blot analysis demonstrated a higher abundance of p-SMAD1/5 in Day 7 blastocysts cultured with miR-148 throughout the entire in vitro culture period (miR148) or from Day 1 to Day 4 of in vitro culture (representing the miRNA effect in the oviduct, miR148-OV). Interestingly, previous studies have reported a decrease in mRNA abundance for SMAD1 and SMAD5 in bovine blastocysts from the 16-cell stage until the blastocyst stage [68], while another study revealed a significant increase in p-SMAD1/5 from the 8–16-cell stage, persisting through Day 7 blastocysts [71]. These findings align with our observations, suggesting a potential explanation for our results. It is plausible that an alternate biological pathway is activated to promote the phosphorylation of SMAD1/5 as a compensatory mechanism. For instance, previous research has shown that BMP2 stimulation induces the phosphorylation of SMAD1/5 in cancer cells [72]. Nonetheless, this hypothesis requires further investigation.
These findings collectively suggest a potential role of miRNA-148b regulating cell proliferation and differentiation in IVP embryos. The importance of miRNAs in maintaining normal organism function raises the possibility that disruptions in maternal miRNA function could contribute to suboptimal embryo development, affecting both natural fertilization and in vitro fertilization programs. However, it is essential to acknowledge that research in this domain faces limitations due to existing technical challenges.
In conclusion, this study demonstrated that miR-148b is taken up by 16-cell embryos and blastocysts when supplemented in the culture medium. This allowed us to assess the effects of miR-148b supplementation in the IVC medium at different time frames, representing its effects in the oviduct or uterus, on embryo quality. We hypothesized that miR-148b could enhance embryo quality and modify the abundance of key genes involved in cell proliferation and differentiation in IVP embryos. These positive responses of miR-148b on embryonic quality in blastocysts produced in vitro could be modulated by BMP/TGF-β through SMAD5 as a possible target gene for miR-148b. Moreover, our results highlight the importance of providing a more comprehensive understanding of the role of the oviduct during early embryogenesis and the potential role of miR-148b in embryo-oviduct interaction.






留言 (0)