
記住我
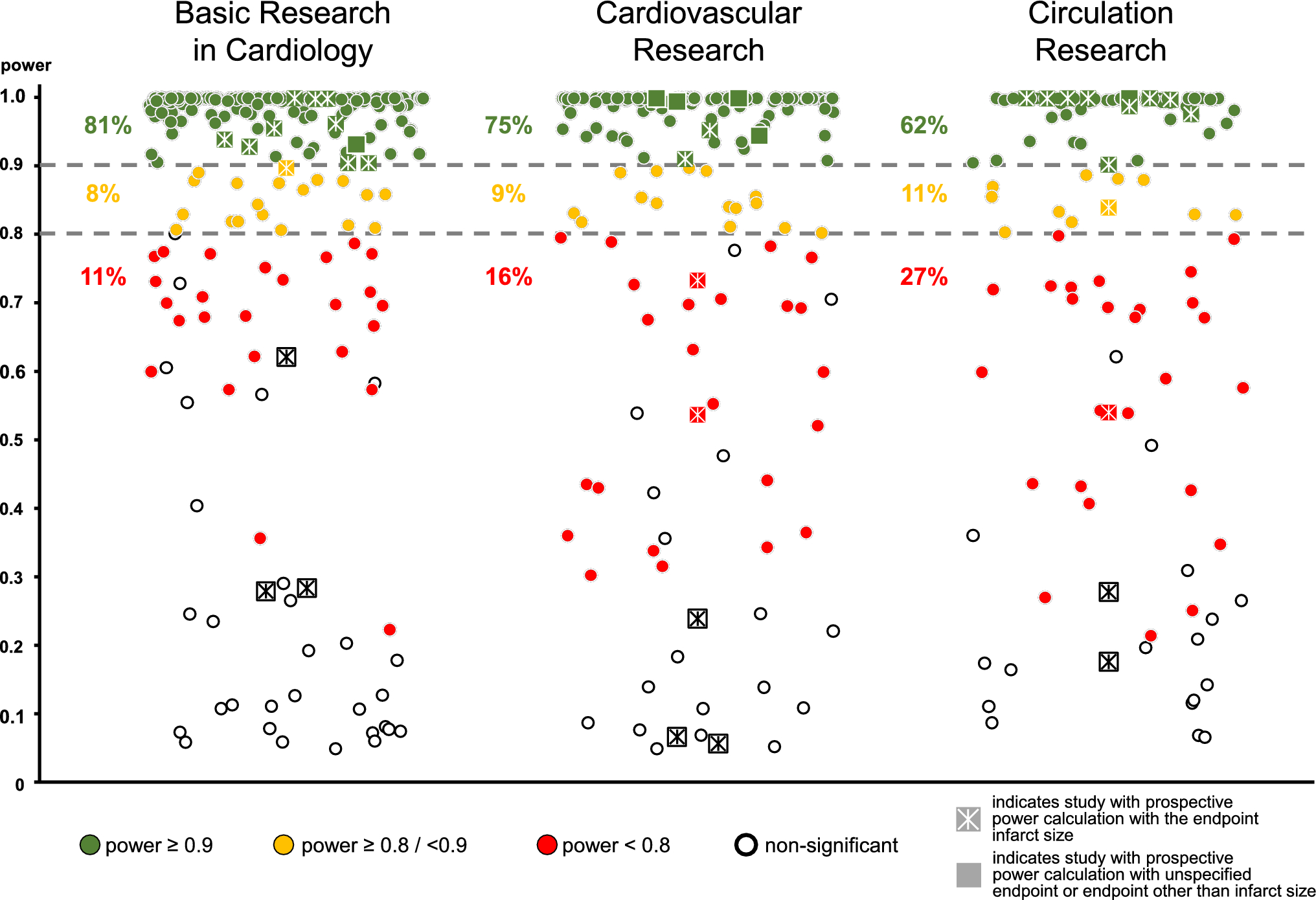


During the first week after MI, the spleen is the major source for monocytes recruited to the myocardium. To replenish the splenic pool splenic myelopoiesis peaks around day 5 [20]. Accordingly, when comparing cell numbers in sham vs MI animals, we found significantly increased monocyte progenitor cells (HSCs and GMPs/CMPs) at day 5 in the spleen but not in the bone marrow (Suppl. Figure 1a,b). Furthermore, the frequencies of EdU+ HSCs, MPPs and GMP/CMPs were significantly higher in the spleens of MI vs sham-operated animals, whereas in the bone marrow the proliferative activity of progenitor cells was unchanged (Suppl. Figure 1c). Moreover, 56 days after MI there were no differences in progenitor cells between sham and MI animals within the spleen and the bone marrow (Suppl. Figure 1a,b).
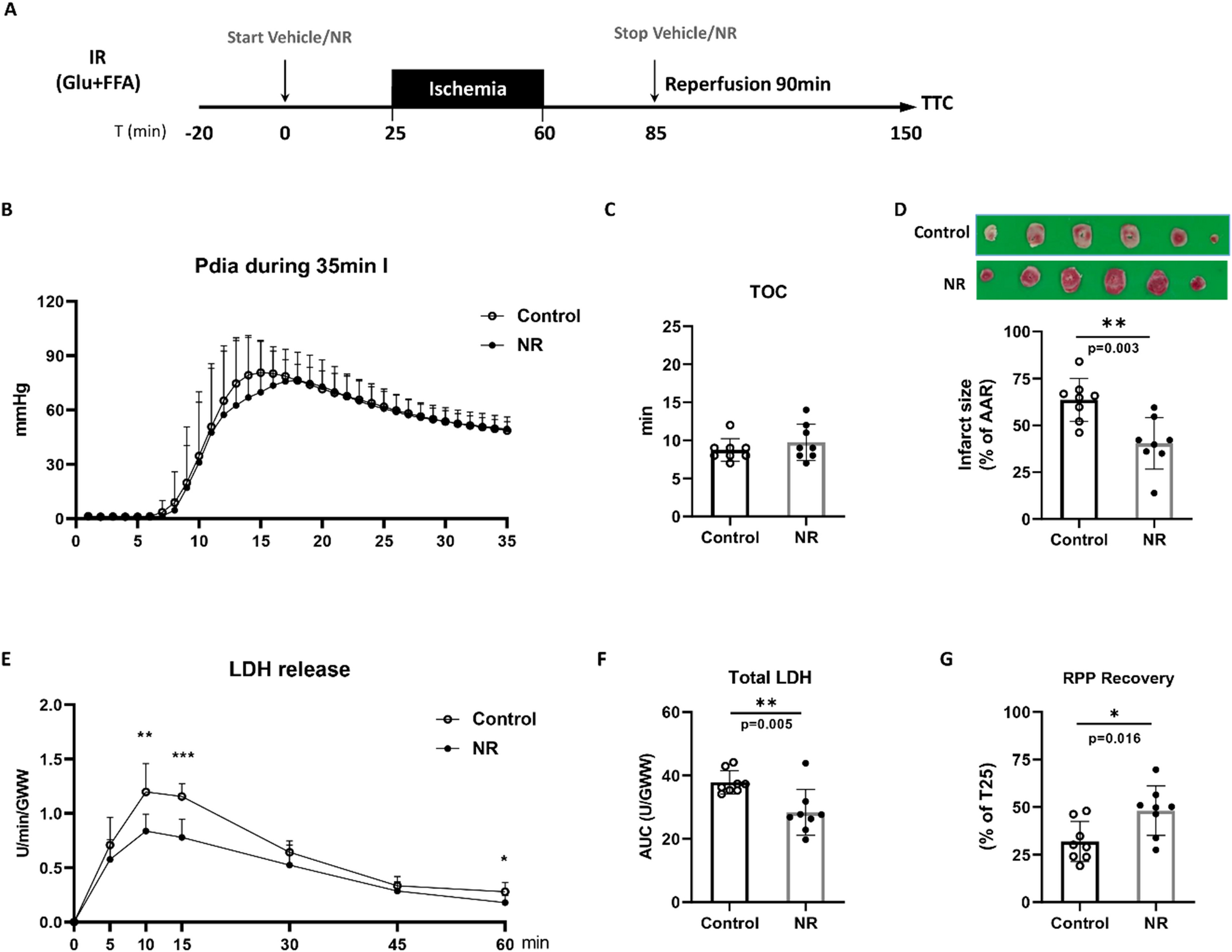
As CD4+ T-cells might play a role in regulating MI-induced splenic-monocyte production, we analysed the composition of the splenic CD4+ T-cell compartment, including both regulatory (Treg: Foxp3+ CD25+) and conventional (Tconv: Foxp3−) T-cells, within the first week after MI (Fig. 1a). The absolute Treg counts on day 5 are significantly higher than on days 3 and 7 after MI. Moreover, there are increased Treg numbers in MI vs. sham on day 5 (Fig. 1b). In contrast, the absolute number of splenic Tconv did not change between days 3, 5, and 7 after MI (Fig. 1c). The Treg:Tconv ratio in the spleen was significantly higher on day 5 when comparing sham vs. MI and compared to day 3 or 7 after MI (Fig. 1d). In comparison to the spleen, no changes in Treg:Tconv ratio between sham and MI were detected in the bone marrow 5 days after surgery (Suppl. Figure 2f). Notably, the frequencies of B- and T-cells are significantly lower in the bone marrow and do not change after MI (Suppl. Figure 2a-e). As these transient changes in CD4+ T-cell composition could affect myelopoietic activity in response to MI, we further studied the role of Tconv and Tregs for myelopoiesis.
Fig. 1
CD4+ T-cell subsets in the spleen. a Flow cytometry plots showing CD25 and Foxp3 expression in splenic CD4+ T-cells. b, c Absolute numbers of CD4+ Foxp3+ CD25+ (Treg; b) and CD4+ Foxp3− (Tconv; c) T-cells in the spleens of post-MI and sham-operated animals on days 3, 5, and 7 after surgery. d Ratio of splenic Treg/Tconv numbers in the infarcted and sham-operated mice 3, 5, and 7 days after sham or MI surgery. MI myocardial infarction. Data are presented as the mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001 (b-d two-way ANOVA)
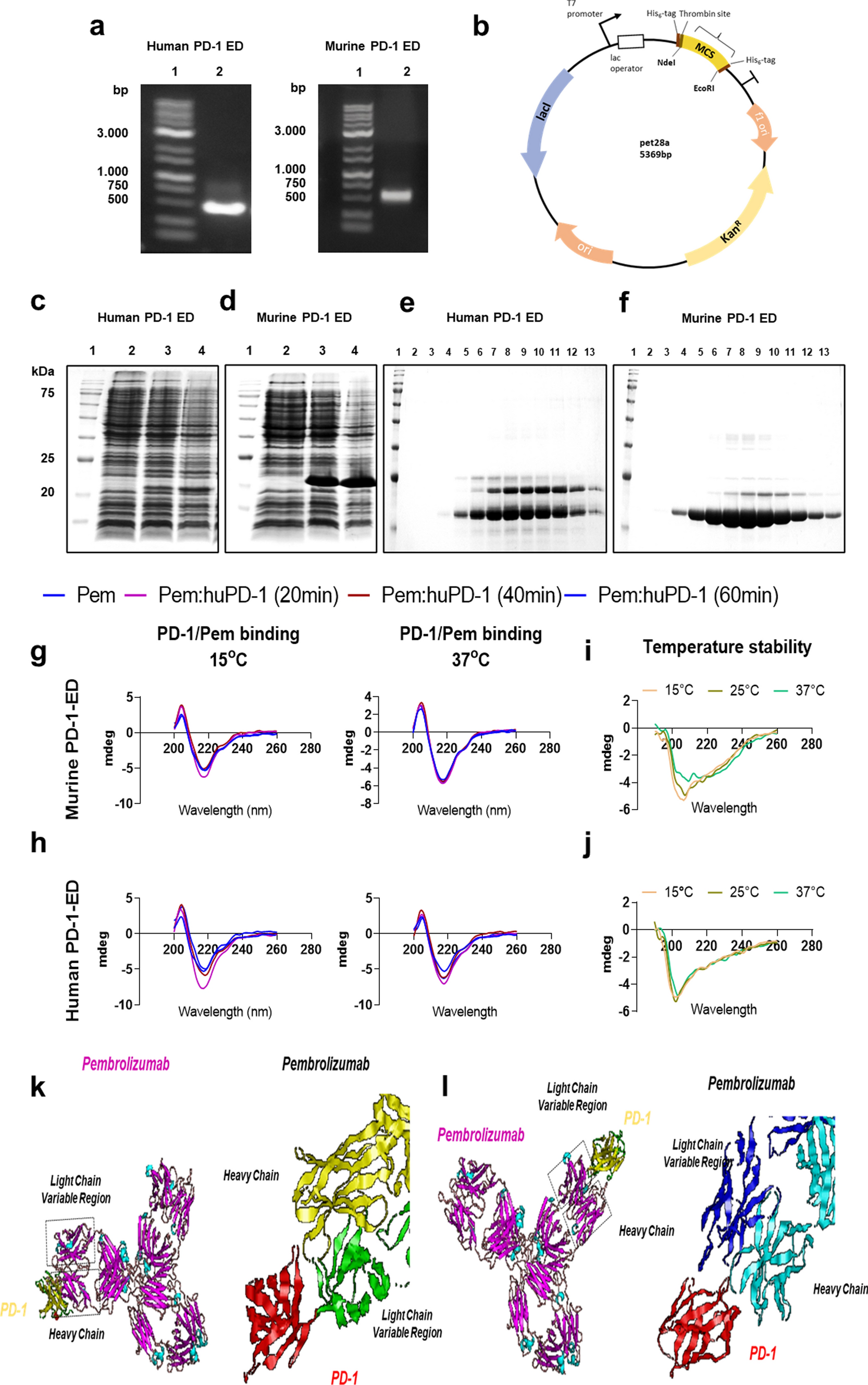
To elucidate the role CD4+ T-cells play in splenic myelopoiesis after MI, we analysed myelopoietic activity in CD4 KO animals vs. WT animals 5 days after MI. We found significantly lower numbers of both myeloid progenitor cells and Ly6Chigh monocytes in the spleens of CD4 KO mice 5 days after MI (Fig. 2a). In contrast to the spleen, no significant differences in progenitor cell numbers and monocytes in the bone marrow of WT and CD4KO mice were found after MI (Fig. 2b). CD4 KO mice show adverse healing with enhanced inflammation in the heart, which might remotely interfere with splenic myelopoiesis [14]. To dissect the impacts of conventional vs. regulatory CD4+ T-cells on myelopoiesis post MI independently from systemic effects caused by different levels of myocardial injury and inflammation, we cultured bone marrow cells from CD4KO mice harvested 5 days after MI. Adding Treg cells did not affect ex vivo proliferation of hematopoietic precursor cells harvested from CD4 KO animals 5 days post MI. In contrast, coculture with activated Tconv enhanced proliferation of precursor cell populations (including HSCs and GMP/CMPs, Fig. 2c). Hence, these results indicate that CD4+ T-cells are required for splenic myelopoiesis in response to MI. Tconv promote myelopoiesis, whereas Treg have no impact in the absence of Tconv.
Fig. 2
Role of CD4+ T-cells in splenic and bone marrow myelopoiesis. Quantification of HSCs, GMPs/CMPs, and monocytes in the spleens a and bone marrow b of WT and CD4 knockout mice (CD4KO) 5 days after MI. c Quantification of proliferative activity of HSCs and GMPs/CMPs isolated from CD4KO BM after coculture with splenic Tregs or Tconv from WT mice. HSC haematopoietic stem cell, GMP granulocyte–macrophage progenitor, CMP common myeloid progenitor. Data are presented as the mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001 (t-test)
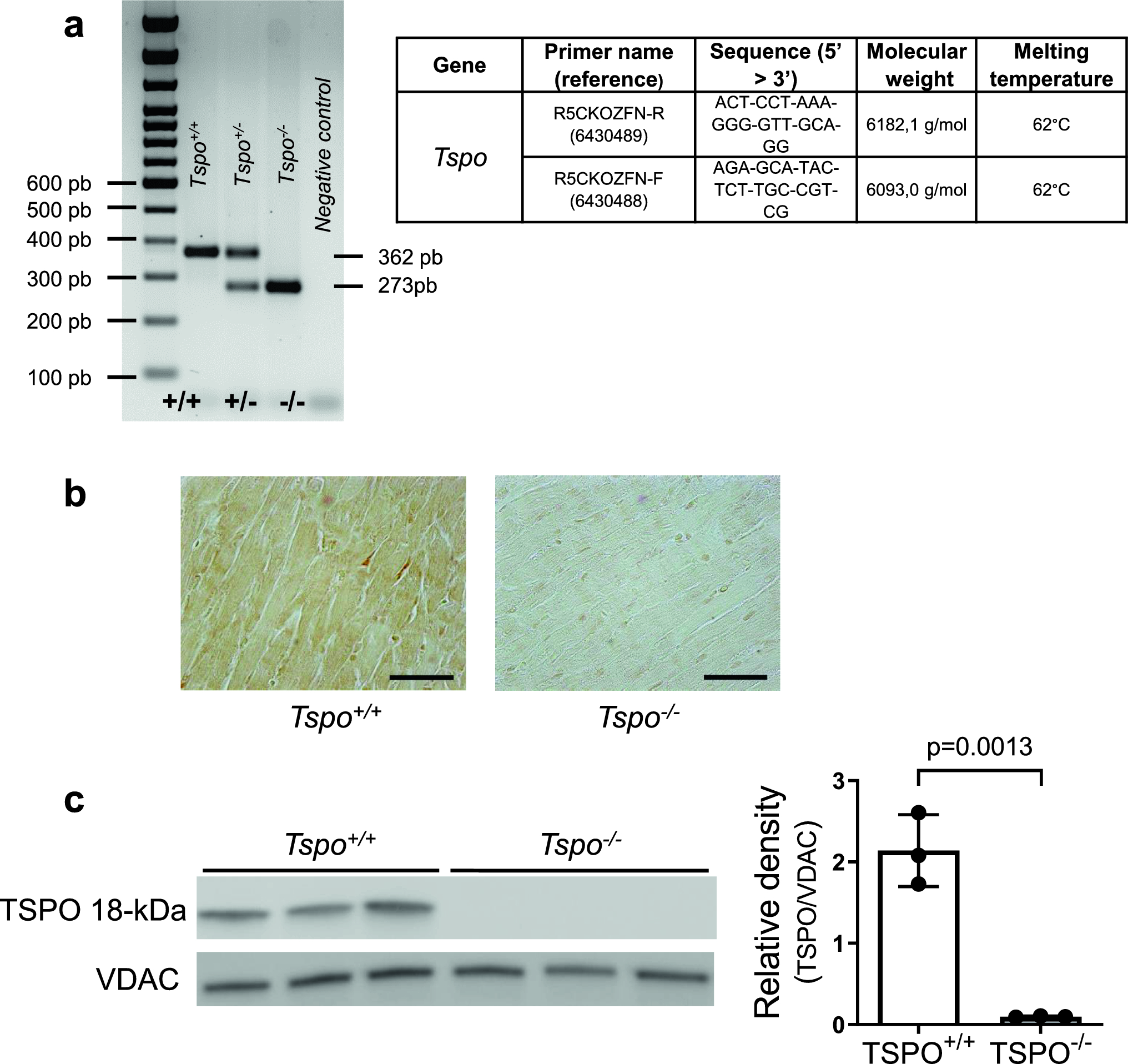
Regulatory T-cell depletion enhances splenic myelopoiesis after myocardial infarctionTo further study the effect of Tregs on splenic myelopoiesis after MI in vivo, we analysed progenitor cell numbers in spleens from WT and Foxp3DTR mice, both treated with diphtheria toxin to selectively deplete Tregs in Foxp3DTR mice (Fig. 3). As we recently showed, this specific Foxp3+ cell depletion approach impairs the clinical outcome after MI by inducing a proinflammatory differentiation state in myocardial macrophages [15]. Notably, our protocol resulted in approximately 60% fewer Foxp3+ cells within the total CD4+ T-cell populations in the secondary lymphoid organs, including the spleen, when comparing Foxp3DTR mice with WT mice (Suppl. Figure 3). Lineage− Sca-1+ CD117+ cell subsets resembling HSCs supplied by the bone marrow and multipotent haematopoietic progenitors (MPPs) and Lineage− CD117± myeloid progenitor cell subsets, including common myeloid progenitors (CMPs), granulocyte–monocyte progenitors (GMPs), common monocyte progenitors (cMoPs), and monocytes, were analysed in the spleen by flow cytometry (Suppl. Figure 4). We studied cellular proliferative activity in vivo by applying the nucleoside analogue 5-ethynyl-2’-deoxyuridine (EdU; Fig. 3a). The absolute number of EdU+ monocytes (Fig. 3b) and the frequency of EdU+ HSCs, MPPs, GMPs/CMPs, cMoPs, and monocytes (Fig. 3c) were significantly higher in infarcted WT mice than in sham-operated WT animals. After MI, the numbers of EdU+ MPPs and cMoPs and the frequencies of EdU+ HSCs, MPPs, GMPs/CMPs, and cMoPs were significantly higher in Foxp3DTR mice than in control WT mice (Fig. 3b, c), further demonstrating the increased in vivo proliferation of progenitor/stem cells in response to infarction in Treg-depleted animals. In the absence of MI, the Foxp3+ T-cell ablation approach per se did not considerably impact splenic progenitor cell proliferation. There was no significant difference in the frequency of EdU+ stem/ progenitor cells between sham-operated WT mice and Foxp3DTR mice on day 5 (Fig. 3c).
Fig. 3
Proliferation analysis and quantification of precursor cells and monocytes in wild type (WT) and Foxp3DTR mice after myocardial infarction. a, b, c In vivo proliferation of HSCs, MPPs, GMPs/CMPs, cMoPs, and monocytes in the spleen 5 days after MI in wild type vs. Foxp3DTR mice. Representative plots a and quantitative data b, c for intracellular EdU analysis in MI animals. d Quantification of HSCs, MPPs, GMPs/CMPs, cMoPs, and monocytes in the spleens of WT and Treg-depleted mice (Foxp3DTR) 5 and 7 days after MI. HSC haematopoietic stem cell, MPP multipotent progenitor; GMP granulocyte–macrophage progenitor, CMP common myeloid progenitor, cMoP common monocyte progenitor. Data are presented as the mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001 (b, c, d two-way ANOVA)
Moreover, the absolute numbers of all hematopoietic stem/ progenitor cell populations per spleen were higher on day 5 than on day 7 after MI in WT mice (Fig. 3d). In Foxp3DTR mice, the total numbers of HSC, MPPs, GMPs/CMPs, and cMoPs were significantly elevated compared to those of WT mice at day 5 after MI (Fig. 3d, Suppl. Figure 5). The number of Ly6Chigh monocytes in the spleen of Foxp3DTR mice remained unchanged compared to that in WT animals after MI (Fig. 3d). However, Ly6Chigh monocytes in diphtheria toxin-treated Foxp3DTR were more numerous in the infarct and border zones of the myocardium and in the blood, compared to those in diphtheria toxin-treated WT mice (Suppl. Figure 6). These comparisons indicate that the increased splenic myelopoietic activity in response to Treg depletion feeds the myocardial monocyte supply, leading to increased proinflammatory monocyte numbers in the myocardium.
Tregs suppress production of the cytokines that drive splenic myelopoiesisNext, we explored whether Tconv in the spleen express cytokines, known to promote myelopoiesis. We detected a transient, modest but significant upregulation of GM-CSF expression in the spleen following MI. GM-CSF expression in the spleen was notably higher in MI vs. sham-operated animals on day 5 (Suppl. Figure 7a). Small subsets of T-cells and B-cells stained positive for intracellular GM-CSF (Suppl. Figure 8a, b) in the spleen, albeit with no difference between the sham-operated and post-MI groups. In addition, we analysed gene expression of other factors that might be involved in splenic Tconv and myelopoiesis crosstalk, including CXCL-12, IL-7, IL-23, IL-2, and IL-1β (Suppl. Figure 7b-f). None of these showed any significantly altered expression in splenic tissue from either sham or MI.
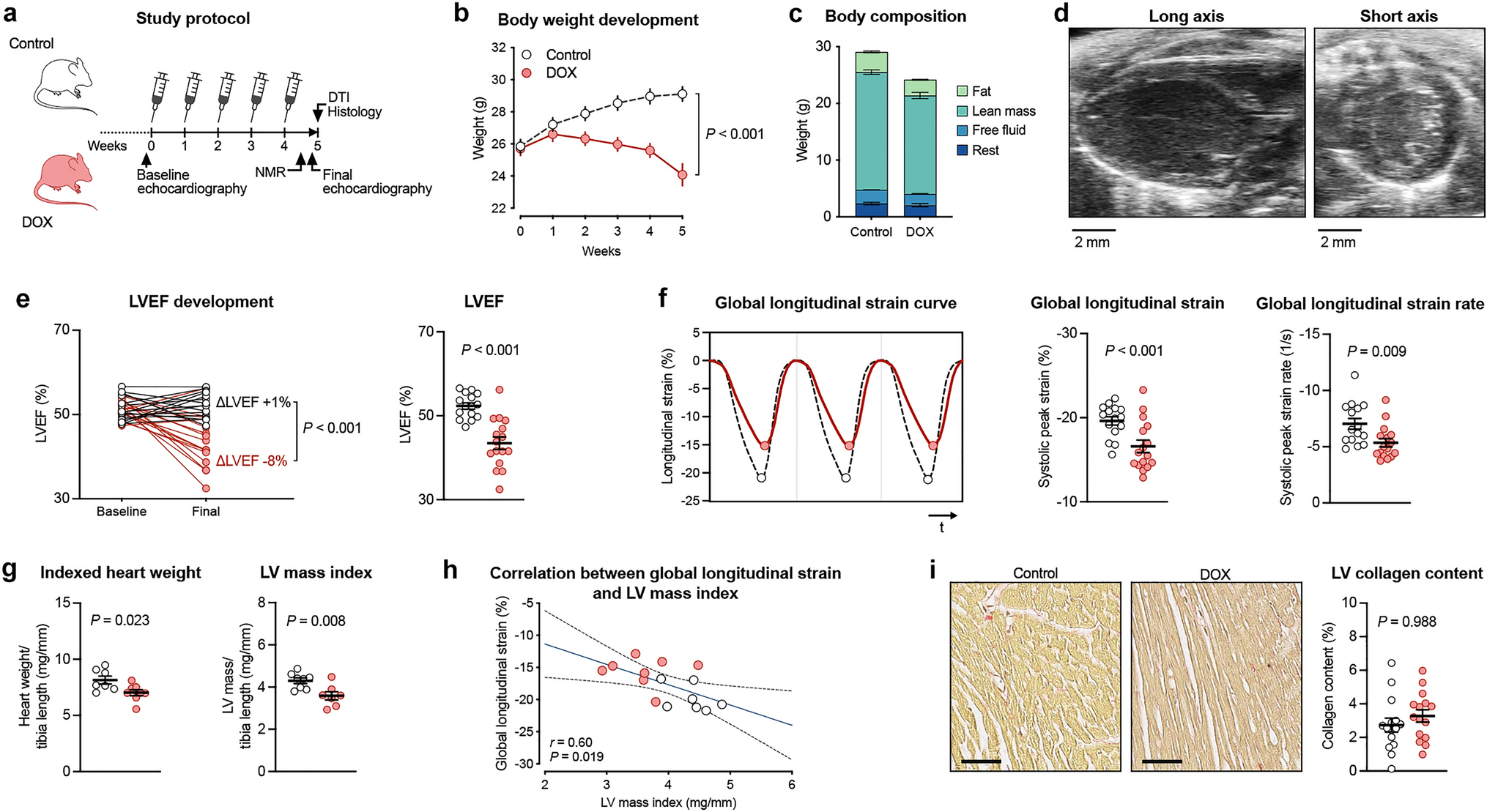
To better understand how Treg–Tconv interaction after MI modulates myelopoiesis, we compared the intracellular expression of those proteins in lymphocytes from WT and Foxp3DTR mice after MI (Fig. 4a). We detected significant increase in the numbers of both IFN-γ- and IL-3-expressing CD4+ T-cells in the spleens of Foxp3DTR mice after MI (Fig. 4b). Furthermore, Foxp3+ T-cell depleted Foxp3DTR mice showed more frequent IFN-γ+ CD4+ and CD8+ T-cells and significantly more frequent IL-3-, GM-CSF, and IL-6-expressing CD4+ T-cells (Fig. 4c). These results indicate that Tregs attenuate the production of several myelopoiesis-promoting cytokines in T-cells.
Fig. 4
Cytokine expression in the spleen 7 days after myocardial infarction in Treg-depleted (Foxp3DTR) and wild type (WT) mice. a Representative flow cytometry plots showing intracellular cytokine expression in CD4+ T-cells of Foxp3DTR and wild type mice 7 days after MI. b, c Expression of GM-CSF, IFN-γ, IL-3, and IL-6 in CD4+ and CD8+ T-cells in the spleens of Foxp3DTR and wild type mice 7 days after MI. c, d Data are shown as the mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001 (t-test)
Treg suppression of splenic myelopoiesis depends on cell-contact and IFN-γTo further mechanistically dissect if Tregs limit myelopoiesis by interacting with Tconvc or through direct effects on progenitor cells, we performed transwell cell culture assays with bone marrow cells from CD4 KO donors. We found HSCs, MPPs, and GMPs/CMPs from CD4 KO donors proliferated more when incubated in a coculture system with Tconv from WT mice, compared to CD4 KO bone marrow cells cultured with Tconv in a transwell system (Fig. 5a, b). This coculture system again underlined the essential role of Tconv in inducing progenitor cell proliferation. Adding Tconv but not Treg cells enhanced proliferation of hematopoietic progenitors. Moreover, the proliferation did not differ in the presence or absence of serum from sham or MI mice, a result that indicates systemic factors released by MI are less important for inducing myelopoiesis (Suppl. Figure 9). Hence, consistent with the results presented in Fig. 2c, Tconv promote progenitor cell proliferation whereas Tregs have no impact on progenitors in the absence of Tconv (Suppl. Figure 9). As IFN-γ represents a prototypical cytokine secreted by activated conventional T-cells, we studied its effect on progenitor cell proliferation in our in vitro system. IFN-γ significantly elevated proliferation of HSCs, GMP/CMPs, and monocytes from CD4 knockout bone marrow (Fig. 6a, b). This indicates IFN-γ has a direct effect on progenitor cells and promotes myelopoiesis in the absence of additional remote factors and CD4+ T-cells.
Fig. 5
Role of Tconv in progenitor cell proliferation in vitro. a Representative flow cytometry plots showing in vitro proliferation of HSCs and GMPs/CMPs from the BM of CD4KO mice in a coculture vs. transwell system with splenic Tconv from WT animals. b The proliferative activity of CD4KO bone marrow precursor cell populations was measured by CFSE staining and normalized to CD4KO BM cultured without splenic Tconv cells from WT animals. Data are presented as the mean ± SD. *P < 0.05, **P < 0.01 (one-way ANOVA)
Fig. 6
Effect of IFN-gamma on progenitor cell proliferation in vitro. a Representative flow cytometry plots showing the proliferative activity of HSCs, GMPs/CMPs, and monocytes, isolated from CD4KO BM, in the presence or absence of IFN-γ. b The proliferative activity was measured by CFSE staining. Data are presented as the mean ± SD. *P < 0.05, **P < 0.01(t-test)
Together, these experiments demonstrate that both cell–cell-contact-dependent mechanism and IFN-γ signalling are vital to the promoting effect CD4+ T-cells have on extramedullary myelopoiesis.
Depletion of Tregs promotes proinflammatory differentiation of monocytesGiven the increased production of proinflammatory cytokines in the spleen in response to Treg depletion, we sought to determine if depleting CD4+ Foxp3+ T-cells might also impact monocyte differentiation in the spleen. Therefore, we sorted Ly6Chigh monocytes from the spleen of WT mice after MI or sham surgery and diphtheria toxin-treated WT and Foxp3DTR mice on day 5 after MI. Next, we performed RNA sequencing for transcriptome analysis (Fig. 7a). In WT animals, MI produced no significant changes in the gene expression of splenic monocytes, compared to animals that underwent sham surgery (Supplementary Table 1 lists the top up- and downregulated transcripts). However, the depletion of Tregs in Foxp3DTR mice led to a significant shift in the transcriptome of splenic monocytes compared to that of monocytes from WT mice after MI (Supplementary Table 2). Notably, the transcriptional regulator Nr4a1, which regulates pro-healing macrophage differentiation from Ly6Chigh monocytes in the infarcted myocardium [13], was amongst the top 50 downregulated transcripts in monocytes from Foxp3DTR mice vs. WT mice after MI. Corresponding to the increased IFN-γ expression in splenic T-cells in Foxp3DTR mice (Fig. 4c), the most significantly upregulated GO set of monocytes from Foxp3DTR mice vs. WT mice after MI relates to transcripts associated with proinflammatory differentiation in response to IFN-γ stimulation (Fig. 7b,c). STRING-based protein–protein interaction network modelling (Fig. 7d) using these top upregulated genes revealed the IFN-γ receptor (Ifngr1) and the IFN-γ-induced chemokine CXCL9 as hubs. We further experimentally confirmed the relevance of IFN-γ for monocyte differentiation upon Treg deletion by analysing the surface-protein abundance of the chemokine receptor CX3CR1, which is amongst the most downregulated transcripts in Foxp3DTR mice (Fig. 7c). Stimulating bone marrow cells ex vivo with IFN-γ downregulated CX3CR1 expression on monocytes (Suppl. Figure 10). Collectively, these data show that Tregs regulate monocyte differentiation in the spleen by mitigating the proinflammatory stimulus imposed by activated Tconv.
Fig. 7
Transcriptome analysis of Ly6Chigh monocytes in the spleens of WT and Foxp3DTR mice 5 days after MI. a After CD3+ and CD19+ cells were depleted by MACS separation columns, Ly6Chigh monocytes were sorted from R1 as CD11b+ Ly6G− F4/80+ Ly6Chigh CD115+ cells. b Gene ontology analysis showing clusters of upregulated genes in Ly6Chigh monocytes of Foxp3DTR vs. those of WT mice. c Cluster analysis of RNA sequencing results depicted as a heatmap (n = 4 animals per group; adjusted p < 0.05 for Foxp3DTR vs. WT). Images were made using the Innate DB database (https://www.innatedb.com/) or the pheatmap package in R (https://www.rdocumentation.org/packages/pheatmap/versions/1.0.12/topics/ pheatmap). d Protein–protein interaction network model of the top upregulated genes. Red represents GO: 0034341“Response to interferone gamma”; Blue represents WP1253 “Type II Interferon signalling” (FDR < 0.0001 each)
留言 (0)