
記住我
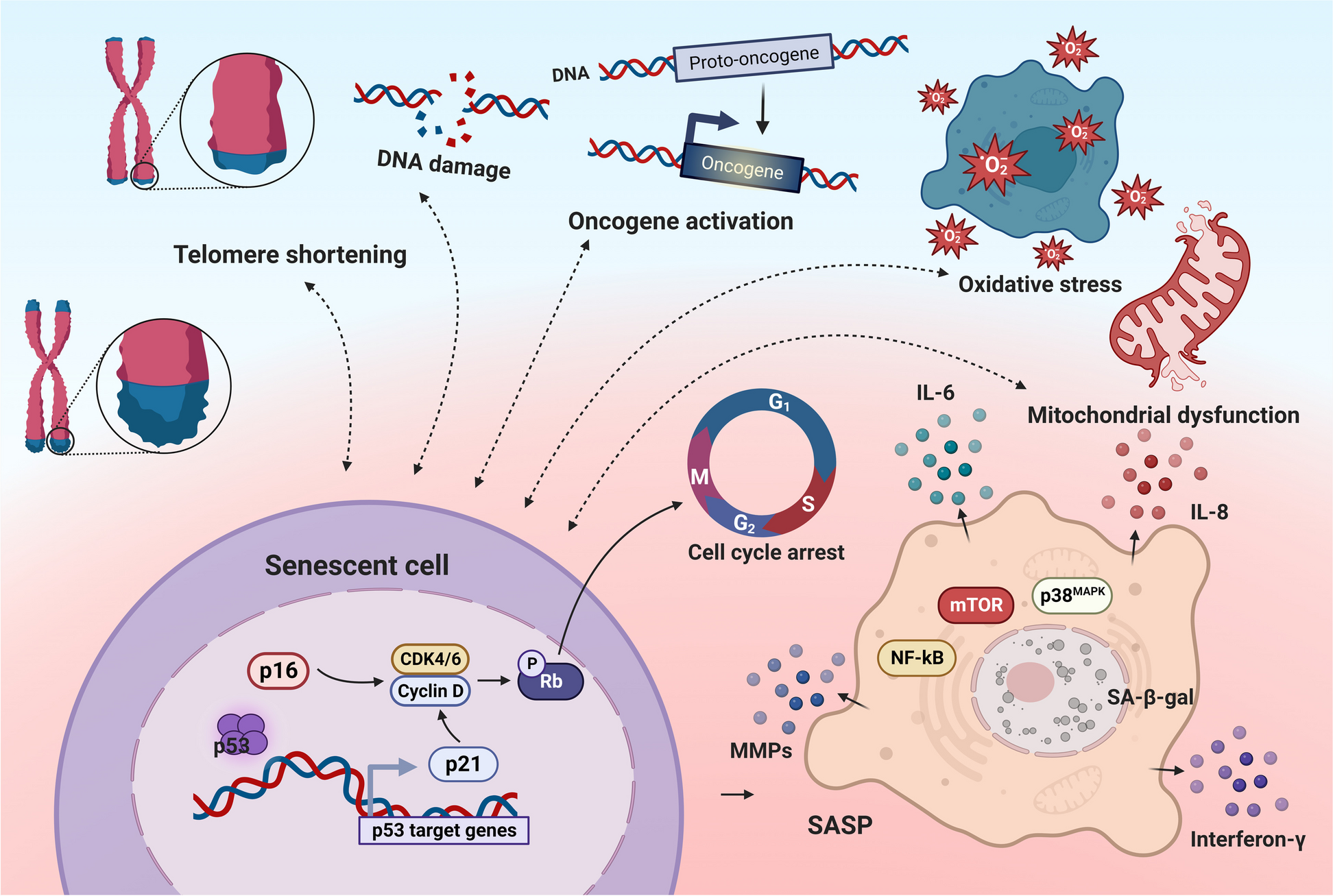
Aging is a complex biological process that affects different organs and tissues in a distinct manner. To discern potential disparities in age-related changes, our investigation embarked on discerning the aging patterns of two distinct systems: the female reproductive system, represented by the ovary and uterus, and the digestive system, represented by the liver. Ovaries, uterus, and livers were collected from C57BL/6 mice at 3-month, 6 months and 9 months of age.
As illustrated in Fig. 1A, the minimal SA-β-gal activity, a well-established marker of cellular senescence [17], was detected in the ovaries of 3- and 6-month-old mice, while a marked increase was observed in the ovaries of 9-month-old mice (Fig. 1B). Additionally, the expression of two pivotal cell cycle suppressor proteins, p16 and p21 [18], were elevated at 6 months and persisting through 9 months of age (Fig. 1C). Conversely, the liver and uterus of 9 months of age, exhibited a limited presence of SA-β-gal-positive cells (Fig. 1D). Apparently, the ovaries exhibited an early and higher accumulation of senescent cells during murine aging.
Fig. 1
Ovary undergoes aging at an earlier stage compared to the liver and uterus. A Representative images of senescence-associated β-galactosidase (SA-β-gal) staining in murine ovarian sections, with corresponding scale bars provided for each image. B Quantitative assessment of the SA-β-gal-positive area in ovarian tissues from 3-month (3 m), 6-month (6 m), and 9-month (9 m) mice (n = 6 for each group). *P < 0.05, **P < 0.01, ***P < 0.001. C Representative immunohistochemical images showing the expression of p16 and p21 in murine ovarian tissues. D Representative SA-β-gal staining images in the uterus and liver sections of mice at 200 × magnification
Age-associated decline in ovarian function correlates with elevated expression of inflammatory factorsAging is charactered by the gradual decline in ovary function. In this study, a substantial increase of TUNEL-positive cells was observed with aging in the ovary of 9-month-old mice, predominantly localized in the ovarian stromal and medullary areas (Fig. 2A), which echoed the areas of SA-β-gal positive staining. Furthermore, the body weight of the three groups of mice increased steadily. Differently, the ovarian weight increased at 6 months but declined at 9 months when compared to the 3-month group, and the ovarian index exhibited a marked decrease at the 9-month (Fig. 2B).
Fig. 2
Age-associated decline in ovarian function correlates with elevated expression of inflammatory factors. A Visualization of TUNEL-labeled apoptotic cells (in green) with DAPI counterstaining (in blue) in the ovaries of mice at 3 months (3 m), 6 months (6 m), and 9 months (9 m) of age. B Comprehensive data on body weight, ovary weight, and ovarian index for each group of mice (n = 12 for each group). C Representative histological images obtained through H&E staining of murine ovarian tissue sections from each experimental group (at 200 × magnification). Atretic follicles are indicated by black arrows. D Quantitative analysis of the average number of follicles per tissue section for each experimental group (n = 5 for each group). E Assessment of relative mRNA expression levels for genes associated with inflammation and senescence-associated secretory phenotype (SASP) (n = 6 for each group). *P < 0.05, **P < 0.01, ***P < 0.001
Ovaries of 3-month-old mice contained abundant numbers of primordial follicles (Fig. 2C). However, at 6 months of age, the number of primordial follicles decreased to approximately 50% of that observed at 3 months, with the most significant decline observed at 9 months (Fig. 2C-D), suggesting a diminish in ovarian reserve during ovarian aging. By histological examination, we observed an increased incidence of atretic follicles and infiltration with inflammatory cells in the 9-month group (Fig. 2D).
The accumulation of senescent cells within ovary is likely to contribute to age-related chronic inflammation [19, 20]. In our study, RT-PCR analysis revealed a significant increased expression of inflammatory molecules (Il6, Il1β, Tnfa, Il18, Il10) in the ovaries of mice, revealing a positive correlation with aging (Fig. 2E). This pro-inflammatory environment should be a significant factor for age-related alterations in ovarian function.
Enhanced oxidative stress and apoptosis in aging granulosa cellsAging exerts multifaceted effects on cellular physiology, with significant consequences observed in various biological systems. Among these, GCs, critical regulators of ovarian function, have been subjected to extensive scrutiny in the context of aging-related changes. In cell aging system induced by H2O2, senescent KGN exhibit marked SA-β-gal activity (Fig. 3A). At the molecular level, there is an upregulation of senescence-associated secretory phenotype (SASP) factors (Fig. 3B), including CDKN1A (protein name: p21), TP53 (protein name: p53), IL6, CXCL8 (protein name: IL-8), IL-1β.
Fig. 3
Enhanced oxidative stress and apoptosis in aging granulosa cells. A Senescent KGN were identified through SA-β-galactosidase (SA-β-gal) staining following treatment with H2O2 (300 µM, 4 h). B Evaluation of relative mRNA expression levels for genes associated with cellular senescence and the senescence-associated secretory phenotype (SASP) (n = 3 for each group). C Intracellular reactive oxygen species (ROS) levels were assessed after H2O2 treatment. Representative fluorescence images depicting aging KGN. D Quantitative analysis of DCFDA staining results (n = 3 for each group). E Measurement of mitochondrial membrane potential (MMP) by JC-10 staining after H2O2 treatment. Representative fluorescence images displaying aging KGN. JC-10 aggregates emit orange-red fluorescence while JC-10 monomers emit green fluorescence. F Flow cytometric assessment of MMP using flow cytometry and presentation of statistical data for MMP analysis (n = 4 for each group). G Annexin V and propidium iodide (PI) staining in KGN treated with H2O2 (300 µM, 4 h). And statistical representation of the apoptosis rate in senescent KGN (n = 4 for each group). Ctrl: control group. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
Additionally, massive ROS production was detected in aged KGN (Fig. 3C-D), suggesting higher oxidative stress in aged KGN. Increased oxidative stress in senescent cells has been linked to the accumulation of dysfunctional mitochondria, manifested by the changes in mitochondrial mass, membrane potential and mitochondrial morphology [21]. As expected, the average mitochondrial membrane potential (MMP) in the aging GCs was significantly lower than that in the control group, indicating that GC senescence was accompanied with mitochondrial dysfunction (Fig. 3E, F). Furthermore, there were more apoptotic KGN in aging group compared with the control group (Fig. 3G).
The ovarian immune milieu is altered with agingThe clearance of these cells represents a crucial process for maintaining homeostasis since the secretion of various cytokines by senescent cells can modify the microenvironment, with the immune system playing a prominent role [22]. We then investigated the disparity of immune cell infiltration and DEGs (Fig. S1A) between murine ovarian tissues at 3 months and 9 months [23]. As showed in Fig. 4A, considerable variations were evident in the composition of immune cells across the tissue samples and age groups in ovarian tissues. Notably, macrophages were the predominant infiltrating cell type (Fig. 4A). Among them, the most frequently observed decline was in M0 macrophages (Fig. 4A). At the protein level, cells expressing F4/80, a well-known macrophage marker [24], were specifically located in the theca of developing follicles at 3 months, which were potentially associated with ovulation. Of note, they were abundant in the interstitial tissue at 6 months (Fig. 4B).
Fig. 4
The ovarian immune milieu is altered with age. A Analysis of immune cell infiltration between ovarian tissues from mice at 9 months (9 m) and 3 months (3 m) of age (Reproduction, 2020). B Immunofluorescence staining of F4/80 in sections of ovarian tissue from mice. C The change of ovarian macrophages polarization over the course of aging. Percentage and number of M1 (CD86 +) and M2 (CD163 +) macrophage from the ovaries of mice at 3 months (3 m), 6 months (6 m), 9 months (9 m) of age were analyzed using flow cytometry (n = 5 for each group). The bar charts reflect the percentage of M1 (CD86 +) and M2 (CD163 +) macrophage within total ovarian macrophages. *P < 0.05, **P < 0.01, ***P < 0.001
Macrophage polarization serves as a candidate biomarker for assessing the inflammatory status and can be roughly classified into two types: pro-inflammatory classically activated macrophages (M1) and anti-inflammatory alternatively activated macrophages (M2) [15]. Here, a combination of CD45, F4/80 and CD11b was used to identify ovarian macrophages (Fig. 4C). Consequently, female mice aged 9 months exhibited elevated levels of the M1 marker (CD86) and reduced expression of the M2 marker (CD163) (Fig. 4C), indicating an age-related alteration in the ovarian immune milieu.
Aging leads to the decline of ovarian macrophages phagocytosisThe DEGs between murine ovarian tissues at 3 months and 9 months were further processed for functional enrichment with Gene Ontology (GO) (Fig. S1B). The results showed that these DEGs are primarily involved in immune responses, including macrophage chemotaxis, cell killing, regulation of inflammatory response, phagocytosis, and macrophage migration. We further demonstrated a significant decrease in the expression levels of phagocytosis-associated molecules, including CD68, CD204, and CD36 in ovarian macrophages of the 9-month group, compared with 3-month group (Fig. S1C, Fig. 5A). Subsequently, the CRA003645 dataset was analyzed for double validation [25]. The heatmap provided a comprehensive overview of DEG patterns within mouse ovarian tissues across three distinct time points: 3 months, 6 months, and 9 months (Fig. S2A). Similarly, GO analysis conducted between 3-month and 9-month group revealed a plethora of DEG associated with phagocytosis (Fig. S2B). Consistent with findings from the 2020 cohort, we observed the decline in M0 macrophages within murine ovarian tissues of the 9-month (Fig. S2C).
Fig. 5
Aging-induced decline in ovarian macrophages phagocytosis. A Flow cytometry analysis was performed to assess the surface expression of scavenger receptors CD36, CD204, and CD68 on macrophages from the ovaries of mice at 3 months (3 m), and 9 months (9 m) of age. The resulting bar charts illustrate the proportion of phagocytic CD36+, CD204+, and CD68+ cells within the total macrophage population (CD11b+/F4/80+), with a sample size of n = 5 for each group. B H2O2-indecued aging macrophages (FITC) were incubated with aging KGN (PE) (at 1:2 ratio) for 3 h. Representative flow cytometry plots was presented and phagocytosis was analyzed by flow cytometry (n = 4, respectively). Ctrl: control group. C Representative flow cytometry plots illustrating the proportions of macrophages within immune cells populations in the follicular fluid (FF) of individuals with diminished ovarian reserve (DOR) and controls (Ctrl). Quantitative analysis of flow cytometry data, presenting the proportion of macrophages (Mφ) within the immune cell population (n = 6, respectively). D Representative flow cytometry plots illustrating the proportions of SA-β-gal+ cells within macrophages in the FF of DOR patients and Ctrl. Additionally, statistical analyses were conducted on these samples (n = 6 for each group). E Representative immunofluorescence images depicting CD68 expression within the FF of DOR and Ctrl individuals. F Gene Ontology (GO) enrichment analysis performed on biological processes between young (control group) and aging primate ovaries macrophages in the dataset GSE130664. *P < 0.05, **P < 0.01, ***P < 0.001
To investigate the influence of aging on macrophage phagocytic activity, H2O2-induced macrophage aging was constructed in vitro, and evidenced by the substantial presence of SA-β-gal-positive cells in the aging group, as well as the elevated levels of SASP factors, including CDKN1A, CDKN2A (protein name: p16), IL6, CXCL8, and IL-1β (Fig. S3A-B). Notably, we observed a pronounced impairment in the phagocytic function of macrophages during aging, as indicated by a substantial reduction in the percentage of engulfed microspheres, declining from approximately 30% to 15% (Fig. S3C). More importantly, when using apoptotic human KGN as targets, a similar decrease in phagocytic capability of aging THP1 cells (human monocytic cell line) was observed (reduced from ~ 20% to ~ 5%) (Fig. 5B). After directly or indirectly co-culturing with aging KGN, there was a significant elevation in the SA-β-gal-positive THP1 in both conditions (Fig. S3D-E), indicating that aging GCs can accelerate the aging process in macrophages.
Subsequently, follicular fluids (FF) were collected from 10 patients with DOR and 10 control subjects. The age of these women ranged from 24 to 35 years, and their serum AMH levels ranged from 0.04 ng/mL to 7.59 ng/mL (Table 1). As shown, a significant decrease in the proportion of macrophages was observed in the DOR group, whereas the SA-β-gal-positive macrophages were notably increased (Fig. 5C, D). Immunostaining with CD68 revealed a less intense signal in the DOR patients compared to the control group (Fig. 5E). Additionally, the single-cell transcriptome data of ovaries from four young monkeys (4–5 years old, 1122 cells) and four aging (18–20 years old, 1479 cells) monkeys [26] were examined. GO analysis on the DEGs between aged and young primate ovarian macrophages demonstrated these DEGs were primarily involved in cell killing, autophagosome assembly, aging, cell migration, and inflammatory response (Fig. 5F).
Table 1 Clinical characteristics of diminished ovarian reserve (DOR)Collectively, the aged ovary exhibited impaired macrophage phagocytosis, likely influenced by aging granulosa cells, indicating intricate cellular dynamics with potential implications for overall ovarian health.
Low molecular weight Chitosan (LMWC) alleviates ovary senescence via promoting macrophages phagocytosisChitosan, a polysaccharide derived from chitin, can be categorized into high, medium, or low molecular weight forms based on its molecular weight range [27]. Previous studies have reported that Chito-oligosaccharides (COS) enhance the phagocytic function of murine peritoneal macrophages following administration through peritoneal injection or intragastric delivery [28]. High-molecular-weight Chitosan (HMWC) has been demonstrated to induce anti-inflammatory polarization, enhance macrophage-released MMP9 activity, and promote migration in human monocyte-derived macrophages [29]. LMWC is distinguished by its heightened permeability, lower melting point, and increased water solubility in comparison to HMWC [30]. However, the specific impact of LMWC on ovarian macrophages and aging remains unexplored.
Cell viability was calculated as previous [31]. Here, we found LMWC significantly enhanced viability in cell proliferation of THP-1 cells, especially at 4 mg/L (Fig. S4A). Treatment with 4 mg/L LMWC significantly increased the expression levels of CD36 and CD204 in PMA-induced M0 cells (Fig. S4B). Moreover, we also investigated in vivo and in vitro the effects of LMWC on macrophage polarization. LMWC treatment significantly enhanced the protein expression levels of CD80, CD163 and CD206 (Fig. S4C-D), inducing M0-like cells to exhibit the characteristics of M1 and M2 macrophages simultaneously. Administration of LMWC significantly increased the mRNA expression of Nos2 (protein name: iNOS) and Il10 (Fig. S4E), suggesting LMWC has a great potential to improve the polarization homeostasis of macrophages. The results of in vivo experiment revealed that LMWC did not affect the estrus cycle (Fig. 6A, Fig. S4F), significantly increased ovary weight and ovarian index (Fig. 6B, Fig. S4G). Remarkably, LMWC administration alleviated SASP-induced DNA damage and cell apoptosis in mouse ovary (Fig. 6C), indicating senescent cells accumulating in aging organs can be effectively cleared through this treatment. Correspondingly, LMWC treatment upregulated the expression levels of CD36, CD68, and CD204 on macrophages in mice (Fig. 6D). Moreover, LMWC-treated mice exhibited a higher count of growing follicles (Fig. 6E). Serum estradiol and AMH levels in the LMWC group were significantly higher compared to the control group, while the levels of FSH levels were lower than that of control (Fig. 6F). The presence of SA-β-gal positive areas in ovarian tissue was notably reduced in the LMWC group (Fig. 6G). These findings suggest that LMWC has the potential to enhance macrophage phagocytosis and phenotypes and further alleviate ovary senescence.
Fig. 6
Low molecular weight Chitosan (LMWC) delays the ovary senescence via promoting phagocytosis of macrophages. A Eight-month-old mice were administrated LMWC in the drinking water (no LMWC as control) for 2 months in prevention experimental settings (n = 10 for each group). At the end point, mice were euthanized with the experimental mice to collect the ovaries for analysis. B Quantification of ovarian index (n = 10 for each group). C Representative images of TUNEL-labeled apoptotic cells (in green) with DAPI counterstaining (in blue) in the ovaries of mice. D Statistical analysis of the proportion of CD36+, CD204+, and CD68+ macrophages within the total macrophages, with a sample size of n = 10 for each group. E Histologic sections of representative ovaries from each treatment group, and average number of the growing follicles per section. F Quantification of serum levels of follicle-stimulating hormone (FSH), estradiol (E2), and anti-Müllerian hormone (AMH) obtained from LMWC-treated and control (Ctrl) mice (n = 10 for each group). G Representative images of senescence-associated β-galactosidase (SA-β-gal) staining in murine ovarian sections, and quantification of SA-β-gal positive area (n = 10 for each group). *P < 0.05, **P < 0.01, ***P < 0.001
留言 (0)