
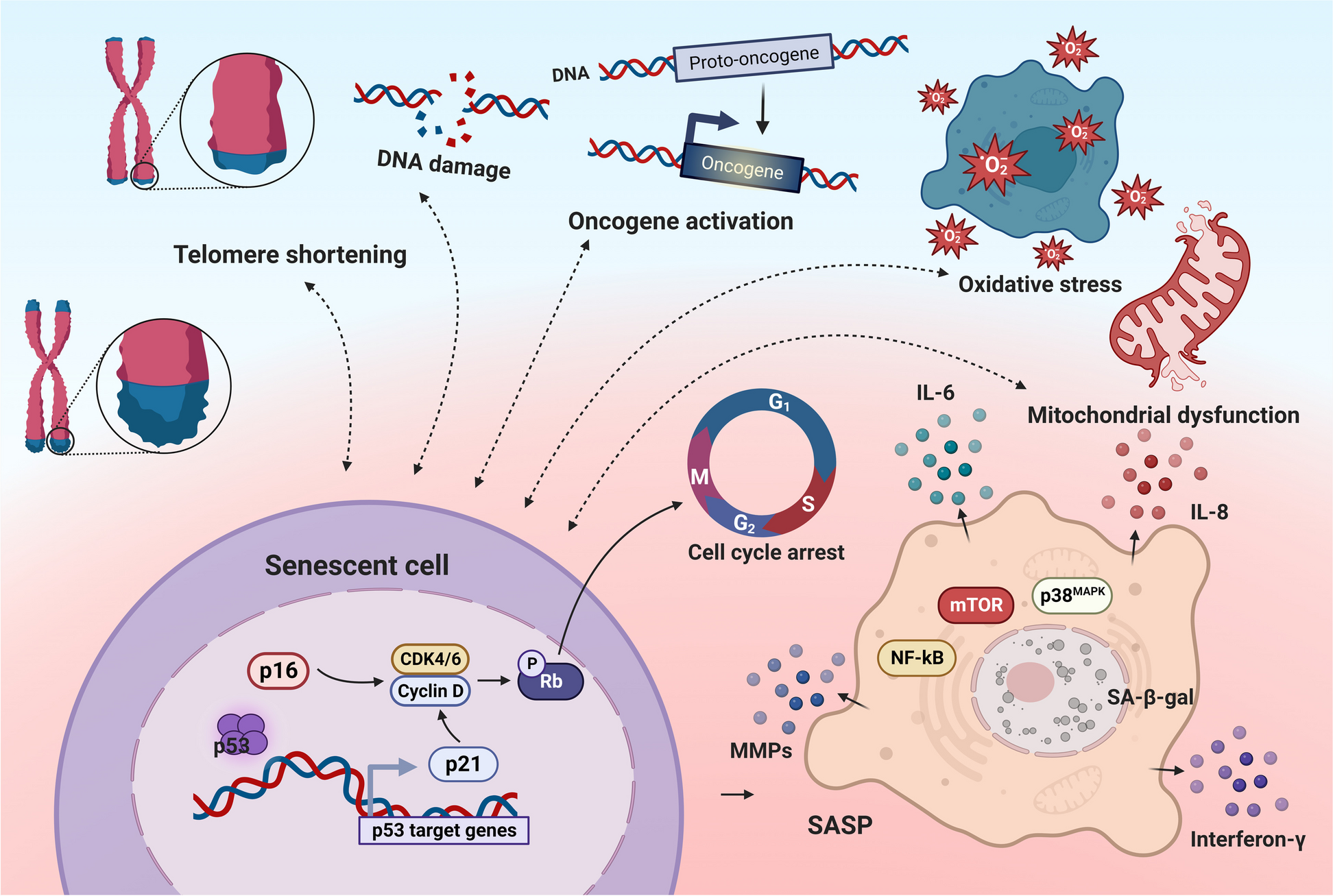
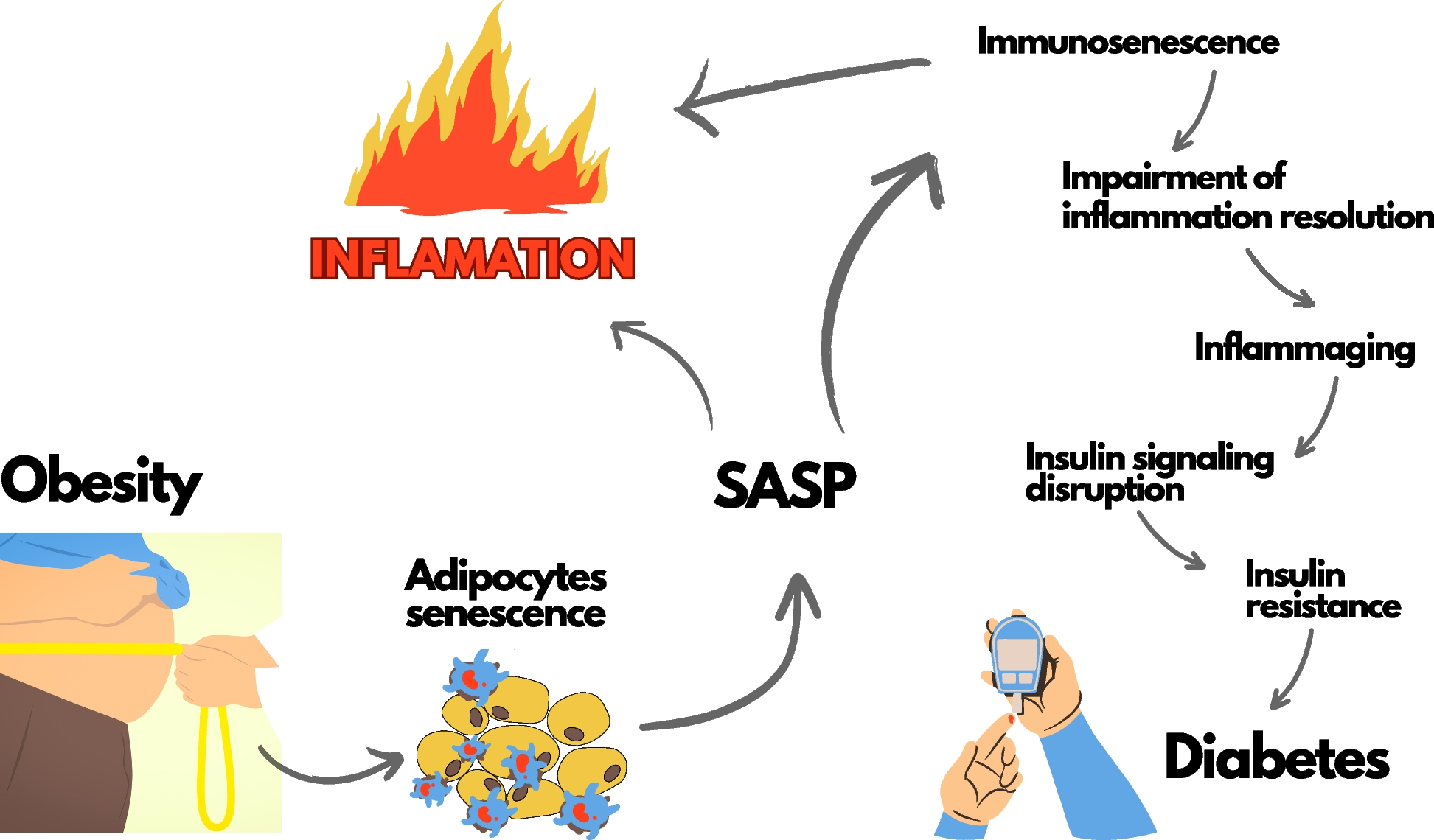
記住我
Total bone marrow was obtained from the femur, tibia, and whole spleen of young and old mice. After digestion and lysis of the erythrocytes, bone marrow and spleen cell suspensions were collected for scRNA-seq (Fig. 1A). After clustering based on previously reported cell markers, all cells were classified into BC, TC, NK, Hematopoietic stem cells (HSC), neutrophil-myeloid progenitor (NMP), DC, monocytes (MONO), macrophages (MAC), neutrophils (NEU), basophils (BASO), red blood cells (RBC), and undefined cells (UNDEF) (Fig. 1B, C and S1A). Compared to young mice, the proportion of cell subgroups in the HIS (comprised of the bone marrow and spleen) of aging mice changed dramatically. With aging, the proportion of lymphocytes decreased markedly, including a pronounced decrease in the proportion of B cells and an increase in the proportion of T and NK cells, whereas the proportion of myeloid cells increased obviously, with an increase in the proportion of BASO, MONO, and NEU, and a decrease in the proportion of MAC (Fig. 1D, E). To further explore the effects of aging, we performed DEG analysis on these cell populations between the 2W and 20M mouse groups and found that cells from old mice expressed higher levels of inflammation-related genes (including the S100 family, AP-1 family, Igha, Ctsg, Ccl5, and Nkg7) and lower levels of growth- and development-related genes (including Sox4, Tgfb1, and Ebf1) than those from young mice (Fig. 1F). In addition, counting the number of upregulated and downregulated DEGs in the cell subgroups revealed a higher number of DEGs in BC, TC, NEU, and MONO, suggesting that aging may have a greater impact on these cell subgroups (Fig. S1B).
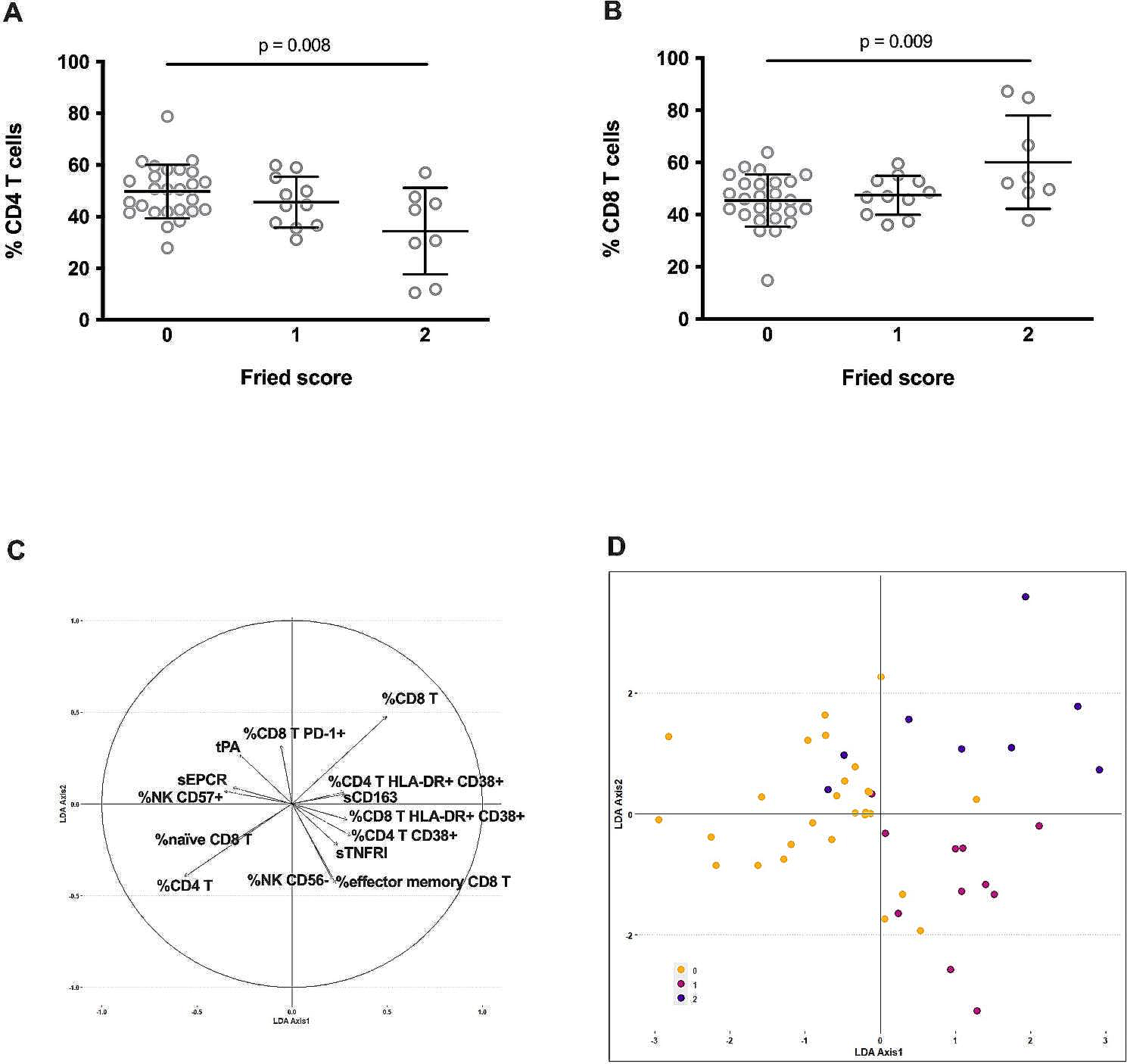
Fig. 1
Overall effects of aging on the characterization of Hematopoietic Immune system. A Schematic diagram of experimental design for scRNA-seq analysis of bone marrow and spleen cells from 2W and 20M mice group. B UMAP plot showing the immune cell types of hematopoietic Immune system in scRNA-seq. C A heatmap showing scaled expression of discriminative gene sets for each cell type. D Bar chart showing the proportion of immune cell types respectively between the two groups of mice. E UMAP plot showing the immune cell types of the hematopoietic immune system in scRNA-seq respectively between the two groups of mice. F Volcano plot showing the up and downregulated DEGs between the two groups of mice. G Representative GO biological processes and pathways enriched in upregulated DEGs in 20M mice. H Representative GO biological processes and pathways enriched in downregulated DEGs in 20M mice. I Violin plot showing the SASP signaling score between the two groups of mice. J Violin plot showing the SASP signaling score among cell types in 20M mice. Significance in I was calculated using the Wilcoxon test; ****P < 0.0001
To better demonstrate the effects of aging on cell function, we conducted enrichment analyses of the upregulated and downregulated DEGs to investigate their biological functions. The results indicated that the upregulated DEGs were mainly enriched in signaling pathways associated with the inflammatory response (such as “inflammatory response,” “immune effector process,” and “positive regulation of cytokine production”) and the immune cell migration and chemotaxis processes (such as “leukocyte migration,” “leukocyte chemotaxis,” and “granulocyte migration”). The downregulated DEGs were mainly enriched in signaling pathways associated with cell growth (such as “chromatin organization,” “protein-DNA complex organization,” and “mRNA metabolic process”) and the cell activation and differentiation processes (such as “B cell activation,” “B cell differentiation,” and “lymphocyte proliferation”) (Fig. 1G, H). The senescence-associated secretory phenotype (SASP) displayed by cells impacts the alterations seen with aging [23]. Therefore, we compared the SASP scores of young and old mice and found that old mice had higher SASP scores than young mice (Fig. 1I). SASP scoring of each cell subgroup in old mice showed that HSC and myeloid cells (including MONO, NEU, and BASO) had higher SASP scores (Fig. 1J). The SASP affects the expression of cytokines and chemokines involved in the inflammatory response. Therefore, we further evaluated and compared the inflammatory response pathway scores between the groups and cell subgroups. We found that the inflammatory response pathway scores of old mice were higher than those of young mice, in which the scores of NEU, MONO, and BASO were higher than those of other cells in old mice, indicating that aging causes the cells in the HIS, especially myeloid cells, to be more involved in inflammatory response pathways, and this may further contribute to aging progression (Fig. S1C, D). Overall, we demonstrated the effects of aging on cell proportions and functions in the HIS and found that aging promoted inflammatory responses and inhibited cell growth and developmental processes.
Aging alters the composition and function of T cells in the HISTo further explore the changes in immune cells during aging, we analyzed cell subgroups. All T cells were clustered and classified into proliferative T cells (PROTC), naïve CD4 + T cells (CD4NA), regulatory T cells (TREG), T helper 1 cells (TH1), T helper 17 cells (TH17), naïve CD8 + T cells (CD8NA), CD8 + effector memory T cells (CD8TEM), CD8 + T cells with cytotoxic activity (CD8CTL), S100 T cells (S100TC), exhausted T cells (TEX), according to cellular markers (Fig. 2A, B). The changes in the proportion of T cells showed a trend consistent with aging. Compared with young mice, old mice had markedly lower proportions of CD4NA, CD8NA, and PROTC, and higher proportions of TEX, TREG, and S100TC. Naïve T cells dominated in young mice, whereas effector and exhausted T cells dominated in old mice (Fig. 2C, D, and S2A). To explore the reasons for this difference in terms of cell development and differentiation processes, we performed a pseudotime analysis in CD4 + T cells and CD8 + T cells. We observed a trajectory from naïve CD4 + T cells to exhausted T cells, and naïve CD8 + T cells to CD8CTL, respectively. Compared with young mice, old mice had more T cells at the end of the trajectory. Specifically, older mice had less CD4NA at the beginning of the trajectory but more TREG and TEX at the end of the trajectory, further indicating that aging maintained more T cells in a highly differentiated state (Fig. 2E, F, and S2B). Six genes with obvious differences in T cells were selected and visualized based on their pseudotimes. Over time, the expression of naïve-related Igfbp4 gradually decreased, whereas the expression of effector and exhaust-related Foxp3 and Tbc1d4 gradually increased. Moreover, the expression of inflammation-related genes Ccl5 and Bhlhe40 increased gradually with time. Among these, Igfbp4 was highly expressed in the young group, whereas other genes were highly expressed in the old group (Fig. 2G and S2C). Enrichment analyses showed that lymphocyte activation and differentiation, adaptive immune response, and inflammatory response processes were upregulated, whereas cell proliferation and differentiation processes were downregulated in the CD4 + T cells of old mice (Fig. 2H and S2D). These findings imply that aging mice are in a state of chronic inflammation involving CD4 + T cells.
Fig. 2
Aging alters the composition and function of T cells in the HIS. A UMAP plot showing the TC subsets of the hematopoietic immune system in scRNA-seq. B A heatmap showing scaled expression of discriminative gene sets for TC subsets. C Pie charts showing the proportion of TC subsets in 2W mice. D Pie charts showing the proportion of TC subsets in 20M mice. E Pseudotime trajectory analysis of CD4 + T cells subsets between two group. Cells are arranged by Pseudotime. F Percentages of CD4 + T cells along the pseudotime for two groups. G Expression transition of Igfbp4, Ccl5, Bhlhe40, Tbc1d4, Stmn1, Foxp3 in CD4 + T cells subsets along the pseudotime. H Representative GO biological process and pathways enriched in upregulated DEGs in CD4 + T cells. I Representative GO biological process and pathways enriched in upregulated DEGs in CD8 + T cells. J Violin plot showing the SASP signaling score between two groups in TC. K Violin plot showing the SASP signaling score among cell types in TC of 20M group. Significance in J was calculated using wilcoxon test; ****P < 0.0001
Pseudotime analysis of CD8 + T cells also showed similar results to CD4 + T cells, with a decreased proportion of naïve CD8 + T cells and an increased proportion of terminal effector CD8 + T cells in old mice (Fig. S2E-G). Continued enrichment analysis of DEGs from CD8 + T cells in old mice showed that these upregulated DEGs were predominantly enriched in pathways associated with inflammatory responses and immune effector processes, whereas the downregulated DEGs were predominantly enriched in biological processes associated with cell proliferation and differentiation (Fig. 2I and S2H). Finally, we used inter-subset SASP scores to investigate the differences in the effects of aging on T cell subsets and found that old mice had higher scores, and PROTC, TH17, and S100TC had the greatest contribution, further demonstrating that aging puts T cells in a highly differentiated state and they become more involved in inflammatory response processes (Fig. 2J-K). In summary, aging increases the number of T cells in a highly differentiated state that are more involved in immune and inflammatory responses against external pathogens.
Aging enhances the immune response of B cells, but reduces the response to new antigensNext, we conducted a systematic analysis of B cells that were classified into six clusters according to specific cellular markers: precursor B cells (PREBC), progenitor B cells (PROBC), naïve B cells (NBC), S100 B cells (S100BC), plasma cells (PBC) and age-associated B cells (ABC) (Fig. 3A-B). A comparison of the proportions of B cell subsets in the two age groups indicated that the proportion of terminally differentiated B cells (ABC, PBC, and S100BC) was 2–3 times higher in old mice than in young mice, whereas the proportion of developing B cells (PREBC, PROBC, and NBC) was significantly reduced (Fig. 3C and S3A). As with the T cells, we performed a pseudotime analysis of these six B cell subsets to explore the effects of aging on their differentiation characteristics over time. Unlike young mice, in which B cells are mostly located at the beginning of the cell trajectory, B cells in old mice are mostly distributed at the end of the cell trajectory. Further visualization of the distribution of B cells with pseudotime showed that more S100BC, ABC, and PBC were present at the end of the cell trajectory in old mice (Fig. 3D-E and S3B). Counting the proportions of cells in each state revealed that old mice had a higher proportion of effector B cells and fewer precursor and naïve B cells at the terminal state-3 of the trajectory (Fig. S3C). Compared to T cells, in young mice, B cells had a more mature functional state, whereas in old mice, B cells contained fewer naïve B cells, indicating a weakened response to new foreign antigens (Fig. 3D, E). In addition, the expression levels of B cell development-related genes (Sox4 and Bcl7a) that were highly expressed in the young group, decreased over time, whereas the expression levels of genes involved in the immune response of B cells (Xbp1, Jchain, S100a6, and Slpi) that were highly expressed in the old mouse group, increased over time (Fig. 3F and S3D).
Fig. 3
Aging enhances the immune response of B cells, but reduces the response to new antigens. A UMAP plot showing the BC subsets of hematopoietic Immune system in scRNA-seq. B The heatmap showing scaled expression of discriminative gene sets for BC subsets. C The dot plot showing the proportion of BC subsets between two groups. D Pseudotime trajectory analysis of BC subsets between two groups. Cells are arranged by Pseudotime. E Percentages of BC along the pseudotime for two groups. F Expression transition of Sox4, Bcl7a, Xbp1, Jchain, S100a6, and Slpi in BC subsets along the pseudotime. G Violin plot showing the SASP signaling score between two groups in BC. H Violin plot showing the SASP signaling score among cell types in TC of 20M group. I UpSet plot showing the integrated comparative analysis of upregulated DEGs in BC subsets. J Representative GO biological process and pathways enriched in upregulated DEGs in BC. Significance in J was calculated using wilcoxon test; ****P < 0.0001
Moreover, the SASP scores of B cells in old mice were higher than those in young mice. Further SASP scoring of B cell subsets in old mice resulted in PROBC having higher SASP scores, indicating that it may be impacted more by aging (Fig. 3G-H). The inflammatory response pathway score for BC in old mice was higher than that in young mice (Fig. S3E). S100BC and ABC showed higher inflammatory response pathway scores in the old mice (Fig. S3F). Next, we sought to determine whether classical inflammatory and immune response-related genes were regulated by aging in a cell type-specific manner. Upset diagrams integrating the up-regulated DEGs in B cell subsets of old mice showed that the inflammatory response-related genes Igha and S100a8 were up-regulated in all B cells [24, 25]. The immune system signaling pathway-related genes Irf8 and Zeb2 [26] were upregulated in ABC (Fig. 3I). Functional enrichment analysis of these upregulated DEGs further supported our hypothesis that aging enables B cells to participate as effector cells in the immune response and drive inflammatory responses (Fig. 3J).
The number and functional status of myeloid cells, especially neutrophils, markedly increases with agingAs identified in the SASP scoring results above, myeloid cells were more affected by aging (Fig. 1J). Genes such as Lrg1, Lcn2, and Prtn3 were upregulated in myeloid cells with aging, and myeloid cells were more involved in multiple immune effector processes as well as leukocyte migration and chemotaxis, whereas genes such as Id2, Jund, and Vcam1 were downregulated, decreasing their involvement in cell proliferation and differentiation (Fig. 4A, B, and S4A). Additionally, the effects of aging on the proportion of myeloid cells should not be ignored. In contrast to lymphocytes, old mice not only had an increased proportion of neutrophils but also an increased proportion of their precursor cells, NMP, indicating a tendency to differentiate into myeloid cells with aging (Fig. 4C and S4B). We further explored the biological functions of the bone marrow cell subsets. We visualized and analyzed subsets of cellular DEGs and found an upregulation of the S100 family and Lcn2 genes in mature myeloid cells. In addition, many genes related to the immune response and cytotoxic molecules (such as Ifitm3, Mpo, and Igkc) were upregulated in the most important NEU subgroup with aging (Fig. S4C). Functional enrichment analysis of myeloid cell subsets with upregulated DEGs showed that NEU and MONO exerted stronger biological functions, including immune responses and bactericidal processes, in old mice (Fig. 4D). Moreover, numerous genes related to mRNA processing and transcriptional activation (Ifg2bp3, Eif5, and Id2) were downregulated in NEU and MONO mice with aging (Fig. S4D). The downregulated DEGs in myeloid cells, especially NEU and MONO, were mostly enriched in cell proliferation- and differentiation-related pathways, such as regulation of the MAPK cascade, chromatin organization, and myeloid cell differentiation (Fig. S4E). These findings indicated that myeloid cells in old mice have a higher differentiation status and play a more robust role in immune effector processes and inflammatory responses. To explore the reasons for these changes, we performed transcription factor analysis on myeloid cells to further investigate the upstream mechanisms. We found that in the top 15 activated TFs, the transcription factors Zfp, Klf10, and Zbtb37 that are associated with RNA polymerase and DNA transcription were elevated in the young mice [27, 28] whereas the immune activation-associated transcription factors Cebpb and hypoxia-inducible factor 1-alpha (Hif1α) were significantly elevated in the old mice (Fig. S4F) [29, 30]. These upregulated TFs and up-regulated DEGs in all myeloid cells identified a potential target gene, Hif1α, that can regulate neutrophil survival through nuclear factor-kappa B (NF-κB) activation and reactive oxygen species (ROS) production (Fig. 4E-G). Also, Hif1α target-gene set scoring of myeloid cells from the two mouse groups further demonstrated a greater effect of aging on Hif1α and its target genes in NEUs (Fig. 4F). Additionally, functional enrichment analysis of up-regulated Hif1α target genes in NEU was conducted to show that the pathway was mainly enriched in neutrophil effector processes involving neutrophil degranulation, immune response, and inflammatory response (Fig. 4G). The inflammatory response process scores of myeloid cells in old mice were higher than those in young mice, with NEUs of old mice having the highest scores, reinforcing previous findings (Fig. S4G-H).
Fig. 4
The number and functional status of myeloid cells, especially neutrophils, markedly increases with aging. A Volcano plot showing the up or downregulated DEGs in MYE. B Representative GO biological process and pathways enriched in upregulated DEGs in MYE. C The pie chart showing the proportion of MYE subsets between two groups. D Representative GO biological process and pathways enriched in upregulated DEGs among MYE subsets. E Venn diagram showing the interaction of upregulated DEGs and upregulated TFs in MYE. F Ridge plots showing the scores of the Hif1a_targeted_ gene_set among MYE subsets between two groups. G Representative GO biological process and pathways enriched in the intersect of Hif1a_targeted_ gene_set and upregulated DEGs in MYE cells. H The flow cytometry histograms (left) and column charts (right) showing the ration of NEU in bone marrow (up) and spleen (down) (n = 5/group). I The flow cytometry histograms (left) and column charts (right) showing the level of Hif1-α in NEU in bone marrow (up) and spleen (down) (n = 5/group). Significance in H-I was calculated using Student’s t test; *P < 0.05, ***P < 0.001, ****P < 0.0001
Consistent with our analysis above, the proportion of NEUs and their expression of Hif1α in the bone marrow and spleen of young and old mice, respectively, was verified by flow cytometry to reveal that there were more NEUs in the old mice and that they expressed higher levels of Hif1α (Fig. 4H-I, S7G). Overall, aging also has a great impact on the proportion and function of myeloid cells in the bone marrow and spleen, especially by increasing the proportion and number of NEU that are involved in immune effector processes and defense against pathogen invasion. Furthermore, we identified the potential target gene Hif1α that regulates neutrophil function after aging and this will help to deepen our understanding of the relationship between aging and neutrophils.
Aging affects the differentiation potential and direction of differentiation of HSCs in the HISAs major immune and hematopoietic organs, the bone marrow and spleen play crucial roles in the production, activation, and differentiation of HSCs. Therefore, it is important to explore the effects of aging on HSCs in the bone marrow and spleen. Volcano plots of DEGs in HSCs from the bone marrow and spleen of young and old mice showed that the upregulated DEGs were mostly related to the inflammatory response and the immune system, whereas the downregulated DEGs were mostly associated with cell activation and differentiation processes (Fig. 5A). Further enrichment analysis of these up- and downregulated DEGs revealed that the upregulated genes were mostly enriched in the biological processes of myeloid cell differentiation, inflammatory response, and immune effector process, whereas the downregulated genes were mostly enriched in the biological processes of chromatin organization, cell activation, and mechanisms associated with pluripotency in the HSCs of aging mice (Fig. 5B and S5A). HSCs in the bone marrow of senescent individuals show a tendency toward myeloid differentiation, and we confirmed this conclusion using HSCs from the bone marrow and spleen. By comparing and fitting the myeloid differentiation and lymphoid differentiation scores of cells in young and old mice, respectively, we showed that the cells in aging mice had a more pronounced tendency for myeloid differentiation (Fig. 5C and S5B-C). Moreover, the SASP score of HSCs in old mice was higher than that in young mice and was impacted more by aging than other cells (except for some myeloid cells), indicating an important effect of aging on HSCs (Fig. 5D and 1J).
Fig. 5
Aging affects the differentiation potential and direction of differentiation of HSCs in the HIS. A Volcano plot showing the up or downregulated DEGs in HSC. B Representative GO biological process and pathways enriched in upregulated DEGs in HSC. C The dot plot showing the lymphoid or myeloid differentiation scores of hematopoietic stem cells, lymphocytes, and myeloid cells. D Violin plot showing the SASP signaling score between two groups in HSC. E Classification of HSC from the three stages into the quiescent phase and other cycling phases (G1, S, and G2M) based on the cell-cycle score of G1/S and G2/M gene sets. F The pie chart showing the proportion of different cell_cycle statuses of HSC between two groups. G The flow cytometry histograms (left) and column charts (right) showing the percentage of LSK CD34 + HSC cells in bone marrow and spleen (n = 5/group). H UMAP plot showing the HSC clusters of hematopoietic Immune system in scRNA-seq (up). The heatmap showing scaled expression of discriminative gene sets for HSC clusters. I The pie chart showing the proportion of HSC clusters between two groups. J. Representative GO biological process and pathways enriched in the discriminative gene sets among HSC clusters. Significance in D was calculated using wilcoxon test; ****P < 0.0001. Significance in G was calculated using Student’s t test; **P < 0.01, ***P < 0.001, ****P < 0.0001
Since HSCs at different developmental stages are a heterogeneous population [31], we performed cell cycle analysis in order to explore the effects of aging on HSCs in the HIS. Cell cycle analysis showed that these stem cells were distributed in the G1, S, and G2/M phases, compared to young mice where HSCs were predominantly clustered in the G1 phase and had a markedly lower proportion of cells in the S phase, indicating that these HSCs were more in the quiescent phase and less activated with increasing age (Fig. 5E-F). Finally, we sampled bone marrow and spleen cells from 2W and 20M mouse groups to validate the effect of aging on the level of function marker in HSCs using flow cytometry. Compared with CD34- HSCs, CD34 + HSCs are in an activated state and exhibit enhanced differentiation and proliferation capabilities. Experimental bone marrow transplantations in both mice and human cells have confirmed that CD34 + HSCs are able to acquire colony formation ability at a faster rate. Flow cytometry results showed that the proportion of CD34 + HSCs in the bone marrow and spleen of 20M mice was lower than that in 2W mice, suggesting that senescence impairs the differentiation and proliferation capabilities of HSCs in the bone marrow and spleen(Fig. 5G and S5D).
To further investigate the mechanisms underlying HSC changes with aging, we classified HSCs into cluster 0 (high-expression markers: Sell, Dntt, and Ccl9), cluster 1 (high-expression markers: Klf1, Tgfbr3, and Gata1), and cluster 2 (high-expression markers: Hlf, Mllt3, and Mecom) (Fig. 5H and S5E). The results of cell ratio analysis and enrichment analysis of DEGs after aging showed that the number of HSC-0 cells that are mainly enriched in the cellular activation and proliferation pathways which means in the hyperactive state, decreased, but the number of HSC-1 cells that are mainly enriched in the myeloid differentiation and proliferation-related pathways, increased (Fig. 5I-J). And this is consistent with our previous speculations. Enrichment analysis of upregulated and downregulated DEGs revealed that after aging, HSC-0 cells were more involved in the inflammatory response and immune effector processes, whereas HSC-2 cells were less involved in the maintenance of cell pluripotency and differentiation processes (Fig. S5G-H).
Given the high response status observed in the HSC-0 subpopulation during functional analysis, we next focused on validating the surface markers and changes of HSC-0 subgroup. We chose the surface marker CD62L (encoded by Sell) for identify this subset. We confirmed that the proportion of HSC-0 (CD62L + HSCs) was decreased in the bone marrow of old mice compared to young mice using flow cytometry. In contrast, this decrease was not observed in the spleen. These findings suggest age-related changes in HSC populations within specific tissue compartments (Fig. S5I). Moreover, CD150 + CD48- HSCs perform a vital function in preserving the long-term differentiation and proliferation abilities of the HSC population [32]. Our study revealed a decline in the population of CD150 + CD48-CD62L + HSCs in both the spleen and bone marrow with aging (Fig. S5J and S6A). CD62L is crucial for cell migration, homing, and steady-state cell proliferation [33, 34]. In previous studies, it has been observed that bone marrow transplantation in mice utilizing CD62L + HSC sorting results in a faster differentiation into terminal lymphocytes and myeloid cells [35]. Similarly, in post-leukemia bone marrow transplantation patients, sorting CD62 + HSC leads to a quicker colonization of bone marrow and differentiation into neutrophils and platelets [36]. Based on these findings, we hypothesize that the CD62L + HSC subpopulation may consist of highly responsive cells that decrease in number with aging.
CD34 + HSCs and CD201 + HSCs demonstrate a more pronounced multi-lineage repopulation capability in immunocompromised mice [37]. We examined and compared the expression of CD34 and CD201 between CD62L + and CD62L- HSC. The finding indicated that the expression ration of CD34 and CD201 in CD62L + HSCs was higher than that of CD62L- HSCs, and we also observed a decrease in CD62L + CD34 + and CD62L + CD201 + HSCs in aging mice, indicating a decline in the high-response function of CD62L + HSCs with age (Fig. S6B-C). In summary, CD62L + HSCs are a rapidly proliferating and differentiating group of high-functioning populations. However, their high-functioning status decreases with aging. And the highly responsive CD62L + HSCs were obviously downregulated in aging, suggesting that they may play an important role in the aging process. Additionally, CD62L plays a crucial role in determining the differentiation fate of common myeloid progenitor (CMP) cells and granulocyte-monocyte progenitor (GMP) cells [38]. In this study, we utilized flow cytometry to analyze the expression of CD62L in CMP and GMP cells during the aging process. Our findings indicate that aging leads to a reduction in CD62L + CMP and GMP cells, suggesting that aging diminishes the differentiation capacity of these progenitor cells (Fig. S6D).
In conclusion, we classified HSCs in the HIS into three subgroups with different functional states and demonstrated the myeloid differentiation tendency and impaired proliferation and differentiation functions of HSC with aging. And CD62L + HSCs may play an important role in age-related diseases as age-related HSCs.
Aging alters intercellular interaction patternsIn the above studies, we found that aging altered the proportion and function of lymphocytes, myeloid cells, and hematopoietic stem cells. However, a simple cell analysis cannot reflect the integral working of the hematopoietic immune system. Therefore, we conducted an intracellular interaction analysis of the hematopoietic immune cells. We analyzed the ligand-receptor (L-R) pairs between hematopoietic immune cells using the CellphoneDB software. Compared to young mice, the interaction between cells in old mice was generally dampened and this was reflected in the decreased communication between myeloid cells, whereas the interaction between DC, T cells, and NK cells heightened (Fig. 6A, B). To explore the specificity of direct cell-to-cell interactions at different ages, we used a Venn diagram to show that L-R pairs were specifically expressed between the two groups. In young mice, the L-R pairs associated with stem cell self-renewal, cell proliferation (KIT_KITLG, FLT1_VEGFB, CCL25_CCR9), and lymphocyte germination (CD40_CD40LG) were highly expressed, whereas in old mice, specific expression of leukocyte adhesion, chemotaxis (SELP_SELPLG), complement secretion (C4A_C5AR2), and inflammatory signals (SELP_CD34) was evident (Fig. 6C). To further explore the different interaction patterns between the young and old groups, we conducted enrichment pathway analysis for genes related to the high expression of L-R pairs in each group. In old mice, pathways related to enhanced immune and inflammatory responses, such as the PI3K-Akt signaling pathway, positive regulation of the ERK1 and ERK2 cascades, and increased secretion of cytokines, were activated (Fig. 6D). Analysis of the expression of inflammatory response-related ligand receptors showed that the interaction between myeloid cells, HSC, and myeloid cells was the most important component in old mice (Fig. S7A). In young mice, the kit receptor signaling pathway was highly enriched, and the pathway related to lymphocyte proliferation and leukocyte differentiation was also highly expressed, and this was also verified by the above findings (Fig. 6E, S7B, and 1H).
Fig. 6
Aging alters intercellular interaction patterns. A The heatmap showing the number of communication interaction L-R among immune cell types in 20M. B The heatmap showing the number of communication interaction L-R among immune cell types in 2W. C Venn diagram showing the interaction of upregulated TFs respectively in two groups. D Representative GO biological process and pathways enriched in the upregulated TF-association genes in 20M. E Representative GO biological process and pathways enriched in the upregulated TF-association genes in 2W. F The dot plot showing the increased L-R signaling of hematopoietic stem cells to myeloid cells in 20M (left) and 2W (right). G The dot plot showing the increased L-R signaling of hematopoietic stem cells to lymphocytes in 20M (left) and 2W (right). H The circle plot showing the MIF signaling pathway network among immune cell types in 20M (up) and 2W (down). I The flow cytometry histograms (left) and column charts (right) showing the level of TNF-α in NEU cells in bone marrow and spleen (n = 5/group). Significance in I was calculated using Student’s t test; **P < 0.01, ***P < 0.001, ****P < 0.0001
CellChat is an important tool for cell-to-cell interaction and pathway visualization. CellChat analysis showed that growth- and development-related pathways (TGFb and BAFF) were upregulated in young mice, while inflammation-related pathways (CD23, CD34, and CD96) were specifically upregulated in old mice (Fig. S7C). To further analyze and verify the relationship between HSCs, lymphocytes, and myeloid cells, we conducted a Cellchat integration analysis in young and old mice. Compared to young mice, the communication probability of L-R associated with inflammation-related MIF was obviously increased in the interaction between HSC and myeloid cells in old mice, while the expression of LGALS9-related L-R pairs was obviously increased in young mice, indicating that the attraction of hematopoietic stem cells to myeloid cells gradually decreases with the progression of aging [39], and this is also consistent with our finding that the proportion of neutrophils in the spleen increased (Fig. 6F, 4H). A similar behavior was observed among HSCs and lymphocytes. In elderly mice, histocompatibility complex-associated ligand receptors were highly expressed in HSC, NK, and T cells, and the expression of macrophage migration inhibitory factor (MIF)-associated L-R pairs increased, whereas in young mice, the L-R pairs related to proliferation and cell aggregation were upregulated (Fig. 6G).
Considering the important role of the MIF pathway, we conducted a visual comparison between young and old mice and found that the interaction between cells increased in old mice and was more significant in myeloid cells, and the genes related to the pathway were also upregulated (Fig. 6H and S7D). At the same time, the TNF signaling pathway [
留言 (0)