
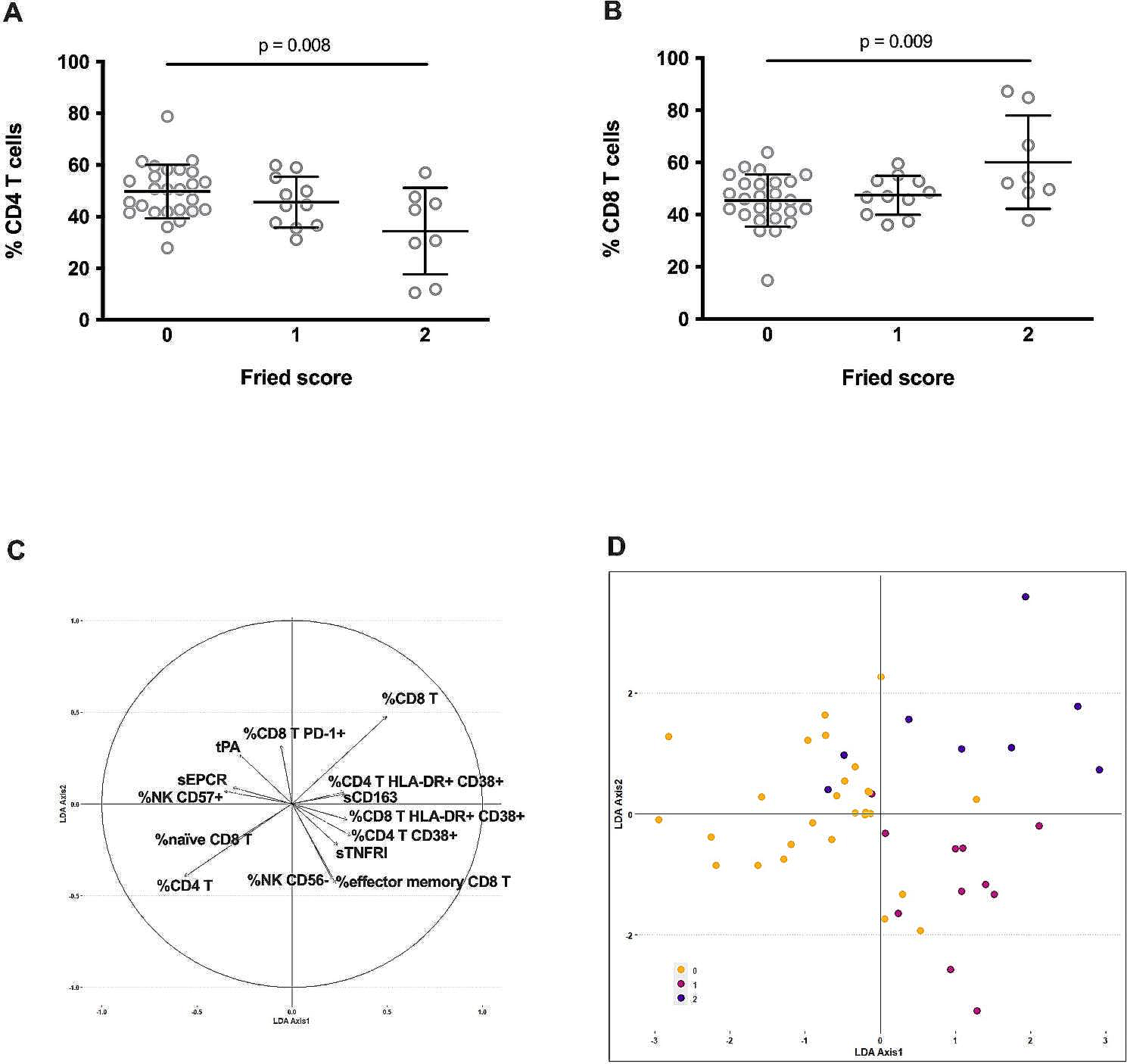
記住我
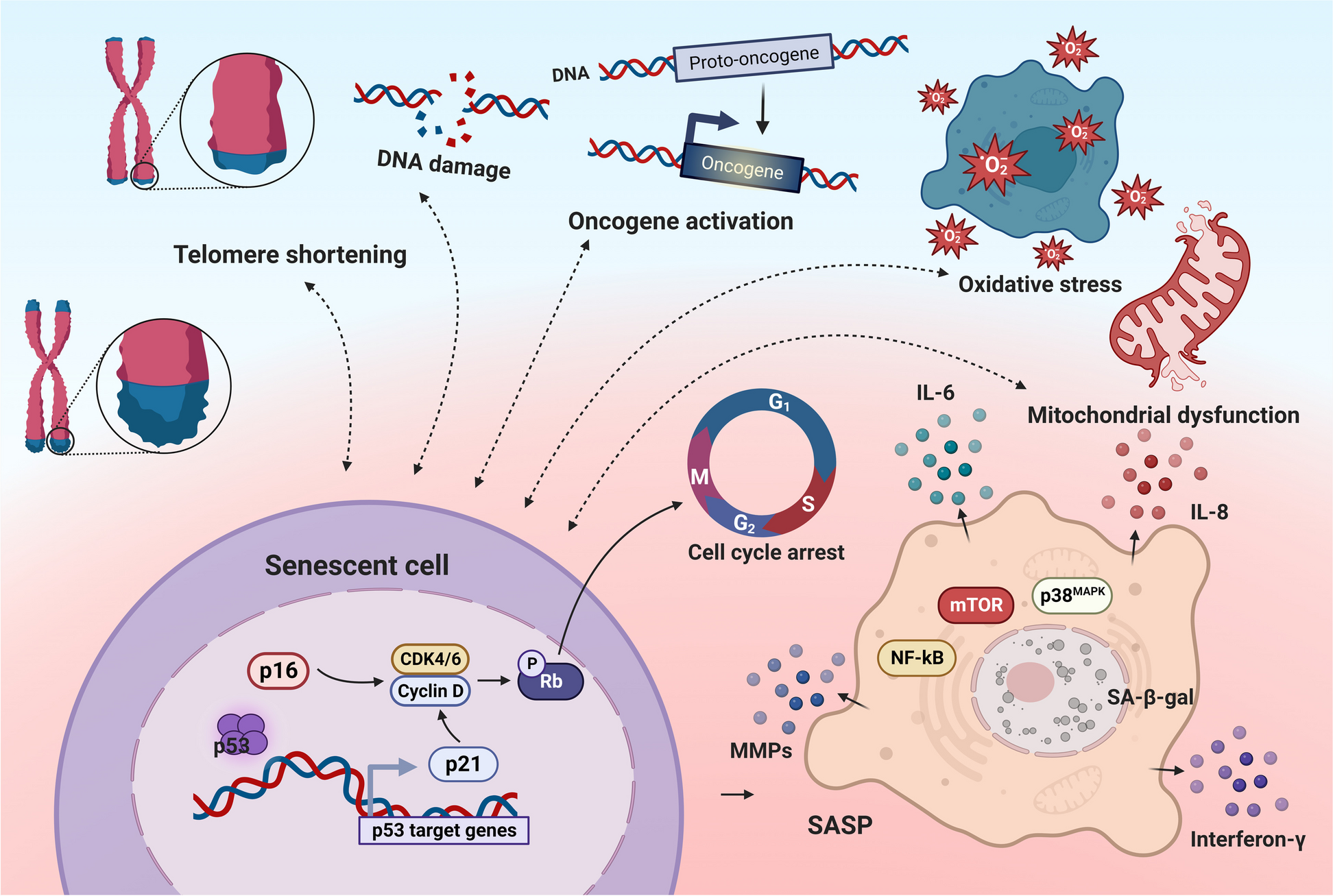
RT‒qPCR was used to analyse the levels of proinflammatory and anti-inflammatory factors in the hippocampus of APP/PS1 mice of different ages (in months) and age-matched WT mice (Supplemental Fig. 1). The mRNA level of IL-17A in the brain increased with age (Fig. 1A). ELISA was used to measure the IL-17A level in the peripheral blood of mice of different ages, and the results were consistent with those observed in the brain (Fig. 1B). Moreover, the mRNA level of TNF-α increased with age, which were consistent with those of previous studies (Fig. 1C). To further explore the effects of IL-17A on cognitive function, cognitive function was assessed using the MWM. We observed that serum IL-17A levels were significantly correlated with escape latency (Fig. 1D).
Fig. 1
Level of IL-17A in samples from model mice and AD patients with different stages of disease. A mRNA levels of IL-17A in the hippocampus of 2-, 5-, and 9-month-old APP/PS1 mice. n = 5 for each group. B Concentrations of serum IL-17A levels in 2-, 5-, and 9-month-old APP/PS1 mice. n = 5 for each group. C mRNA levels of TNF-α in the hippocampus of 2-, 5-, and 9-month-old APP/PS1 mice. n = 5 for each group. D Relationship of serum IL-17A concentration with escape latency in 9-month-old APP/PS1 mice. E Serum IL-17A levels in AD patients with different stages of disease. F Serum IL-17A levels in patients with AD and other neurodegenerative diseases. G Relationship of the serum IL-17A level with the MoCA score of AD patients according to linear regression analysis. H Relationship of the serum IL-17A level with the MMSE score of AD patients according to linear regression analysis. One-way analysis of variance was used to assess the differences among more than two groups. The values are expressed as the mean ± standard deviation (****P < 0.0001, ***P < 0.001, **P < 0.01, and *P < 0.05). HC: healthy controls; MCI: mild cognitive impairment; DAT: dementia of Alzheimer type; VaD: vascular dementia; PD: Parkinson’s disease. The data are representative of at least three independent experiments
IL-17A in AD patients at different stagesBlood samples were collected from patients with neurodegenerative diseases (including AD, PD, and VaD), and sex- and age-matched HC were recruited from among physical examination patients. Figure 1E shows the concentrations of IL-17A in the samples from AD patients with different degrees of disease progression. The detectable levels of IL-17A were significantly higher in AD patients than in the controls. Moreover, the serum levels of IL-17A in PD and VaD patients were also higher than those in the controls (Fig. 1F). Linear regression analysis showed that the serum IL-17A level was significantly correlated with the MMSE and MoCA scores of AD patients (Fig. 1G-H). These results indicate that IL-17A may play a role in the progression of AD and is associated with cognitive function.
IL-17A exacerbated cognitive impairment in APP/PS1 miceBased on the above studies, the levels of IL-17A increased with the age of APP/PS1 mice. To further explore the effects of IL-17A on cognitive function, we intraperitoneally injected APP/PS1 mice with recombinant IL-17A or anti-IL-17A using PBS and isotype (ISO) as controls (Fig. 2A). Cognitive ability was assessed by the MWM test. As shown by the MWM tests, compared with the injection of PBS, injection of IL-17A significantly decreased cognitive function of the APP/PS1 mice, and their escape latency significantly increased on day 5. Compared with the injection of ISO, the injection of anti-IL-17A substantially improved the cognitive function of APP/PS1 mice. The escape latency of the APP/PS1 mice was markedly reduced from day 2 to day 5 (Fig. 2B). Compared with the PBS injection, the Aβ1-40 in the brain tissue of mice considerably increased after the injection of recombinant IL-17A, and Aβ1-42 showed no sizable change (Fig. 2C–D). Subsequently, we performed thioflavin-S staining to examine senile plaque deposition in the brain tissues of the mice. After the intraperitoneal injection of recombinant IL-17A, 9-month-old APP/PS1 mice exhibited an increase in senile plaque deposition in brain tissues, which was effectively reduced by anti-IL-17A antibody treatment (Fig. 2E). Altogether, these data suggest that IL-17A substantially accelerates Aβ plaque deposition in the brain tissue of APP/PS1 mice and that anti-IL-17A treatment results in a decrease in Aβ plaque deposition.
Fig. 2
IL-17Ab ameliorated the decrease in spatial and working memory in APP/PS1 mice. A Schematic of IL-17A or IL-17Ab administration to APP/PS1 mice via intraperitoneal injection. B Escape latency of APP/PS1 mice injected with IL-17A or IL-17Ab during the training phase of the MWM test. n = 5 for each group. C Aβ1-40 levels in the brain tissues of APP/PS1 mice after the injection of IL-17A or IL-17Ab. D Aβ1-42 levels in the brain tissues of APP/PS1 mice after the injection of IL-17A or IL-17Ab. E Representative immunofluorescence images of Aβ plaques in APP/PS1 mice after the injection of IL-17A or IL-17Ab. The values are expressed as the mean ± standard deviation (****P < 0.0001, ***P < 0.001, **P < 0.01, and *P < 0.05). The data are representative of at least three independent experiments
IL-17A elevated the level of inflammation via the TLR4/NF-κB signalling pathway and microglial activation in the brains of APP/PS1 miceThe above studies showed that after the injection of IL-17A in vitro, the cognitive function of APP/PS1 mice decreased, the escape latency was prolonged, and Aβ plaques were deposited in the brain. However, the IL-17A antibody can considerably improve the cognitive function of APP/PS1 mice. We speculate that IL-17A increases neuroinflammation by changing the levels of inflammatory factors in the brain, thus exacerbating cognitive decline. We observed that the trend of change in TNF-α in the brains of APP/PS1 mice was consistent with that of IL-17A. Thus, we speculated that IL-17A injection in vitro can increase the level of TNF-α in the brain.
The hippocampal tissues of the above model mice were collected, and the TNF-α levels in the hippocampi were measured by RT‒qPCR and ELISA. The results showed that the mRNA expression of TNF-α in the brain tissue substantially increased after the injection of recombinant IL-17A, and this expression was effectively reduced by anti-IL-17A treatment compared with ISO (Fig. 3A). TNF-α levels were measured by ELISA and found to be consistent with the mRNA levels (Fig. 3B). These results indicate that IL-17A may affect the progression of AD by regulating TNF-α.
Fig. 3
IL-17A elevated the level of inflammation through the TLR4/NF-κB signalling pathway and microglial activation. A TNF-α mRNA levels in the brain tissue of APP/PS1 mice after injection of IL-17A or IL-17Ab were determined by RT‒qPCR. B TNF-α levels in the brain tissue of APP/PS1 mice after the injection of IL-17A or IL-17Ab were determined by ELISA. C Expression levels of TLR4/NF-κB signalling pathway-related proteins in the brain tissues of APP/PS1 mice that were injected with IL-17A or IL-17Ab. D–F mRNA expression levels of TLR4, NF-κB, and MYD88 in the brain tissue of APP/PS1 mice after the injection of IL-17A or IL-17Ab. G Representative immunofluorescence images of IBA-1+ microglia in APP/PS1 mice after the injection of IL-17A or IL-17Ab. The values are expressed as the mean ± standard deviation (****P < 0.0001, ***P < 0.001, **P < 0.01, and *P < 0.05). The data are representative of at least three independent experiments
Microglia express a variety of receptors, including TLRs, which are pattern recognition receptors [15]. When microglia are stimulated by debris from apoptotic and necrotic cells and heat shock proteins, TLR4 is activated, which further stimulates the downstream transcription factor NF-κB and promotes the secretion of the proinflammatory cytokines TNF-α, IL-1β, and IL-6 [16]. IL-17A interacts toward specific receptors (IL-17RA and IL17RC). This interaction leads to a cascade of intracellular signaling pathways that induces direct and indirect activation of the NF-κB signaling pathway. The expression levels of IL-17RA, IL17RC and key proteins in the TLR4/NF-κB signalling pathway in the brains of the model mice were measured. Considering the activation of TLR4, the expression of CD80, CD86 and MHCII were also detected. WB data revealed that the APP/PS1 mice that were treated with recombinant IL-17A exhibited increases in the expression of TLR4, MYD88, NF-κB, IL-17RA, IL17RC, CD80, CD86 and MHCII compared with the control mice. Anti-IL-17A antibody treatment caused the downregulation of TLR4, MYD88, NF-κB, IL-17RA, IL17RC, CD80, CD86 and MHCII in the hippocampal tissues of APP/PS1 mice (Fig. 3C, Fig. S2). The mRNA levels were consistent with the protein levels (Fig. 3D–F). These results suggest that IL-17A may enhance TNF-α levels in the brain by activating the TLR4/NF-κB signalling pathway.
Microglial activation is a part of the immune response of the human brain [17]. We used IBA-1 as a specific marker to analyse the changes the microglia of the cortex and hippocampus of APP/PS1 mice that were treated (8 months) with recombinant IL-17A or an anti-IL-17A antibody. In contrast to the activated microglia (amoeboid) that were observed in APP/PS1 mice that were treated with PBS, increased microglial accumulation was observed in the cortex and hippocampus of APP/PS1 mice that were treated with recombinant IL-17A. However, the number of IBA-1-positive microglia markedly decreased after the injection of the anti-IL-17A antibody (Fig. 3G). Thus, our results indicate that IL-17A activates microglia, which secrete inflammatory mediators (TNF-α) to induce neurotoxicity.
IL-17A upregulated TNF-α through the TLR4/NF-κB signalling pathway in BV2 cellsBy establishing an animal model of IL-17A overexpression, we discovered that IL-17A increases the level of TNF-α in the brain through the TLR4/NF-κB signalling pathway, aggravates neuroinflammation, and thus exacerbates cognitive impairment. The BV2 cell line is a type of microglial cell line derived from C57/BL6 mice. BV2 cells were used as model cells to further verify the role of IL-17A in neuroinflammation at the cellular level. BV2 cells were stimulated with lipopolysaccharide (LPS) (100 ng/mL) for 0, 4, 8, and 24 h. The culture medium was collected, and the TNF-α level in the culture medium was measured by ELISA. The results showed that the level of TNF-α in the culture medium was significantly higher than that in the control group after 24 h of stimulation (Fig. 4A). BV2 cells were collected, and the expression level of TNF-α mRNA was measured by RT‒qPCR. The results showed that the TNF-α mRNA level in BV2 cells markedly increased after stimulation with LPS for 4 h (Fig. 4B). These results suggest that microglial secretion of TNF-α considerably increases under inflammatory conditions.
Fig. 4
IL-17A upregulated TNF-α through the TLR4/NF-κB signalling pathway in BV2 cells. A Changes in the TNF-α levels the supernatants of BV2 cells that were stimulated with LPS at different times (ELISA). B Changes in TNF-α mRNA expression in BV2 cells that were incubated with LPS at different times (RT‒qPCR). C TNF-α levels in the supernatants of BV2 cells that were incubated with LPS and IL-17A for 24 h. D TNF-α mRNA levels in BV2 cells that were incubated with LPS and IL-17A for 4 h. E Expression levels of TLR4/NF-κB signalling pathway-related proteins in BV2 cells after IL-17A stimulation for 24 h under inflammatory conditions. F–H mRNA expression levels of TLR4, NF-κB, and MYD88 in BV2 cells that were stimulated with IL-17A for 24 h under inflammatory conditions. The values are expressed as the mean ± standard deviation (****P < 0.0001, ***P < 0.001, **P < 0.01, and *P < 0.05). The data are representative of at least three independent experiments
We further explored the effect of IL-17A on TNF-α secretion by BV2 cells. The ELISA results showed that the TNF-α levels in the culture medium of BV2 cells that were incubated with IL-17A (20 ng/ml) and LPS for 24 h was considerably higher than those of cells that were incubated with LPS and IL-17A alone (Fig. 4C). The RT‒qPCR results showed that the expression level of TNF-α mRNA in BV2 cells was markedly higher than that in the LPS and IL-17A alone groups after 4 h of coincubation with IL-17A and LPS (Fig. 4D). These results indicate that IL-17A further promotes the synthesis and secretion of TNF-α in microglia during neuroinflammation. In addition, WB or RT‒qPCR were used to measure the expression of IL-17RA, IL17RC, CD80, CD86, MHCII and related proteins in the TLR4/NF-κB signalling pathway. We observed that IL-17A readily upregulated the levels of TLR4, NF-κB, and MYD88 transcripts and stimulated the production of TLR4, NF-κB, MYD88, IL-17RA, IL17RC, CD80, CD86 and MHCII proteins in vitro (Fig. 4E–H, Fig. S3), further confirming that IL-17A increases the TNF-α levels by activating the TLR4/NF-κB signalling pathway.
IL-17A decreased the activity and phagocytosis of BV2 cellsThrough the above studies, we demonstrated that at the cellular level, IL-17A may increase TNF-α secretion by microglia via the TLR4/NF-κB signalling pathway. We then aimed to elucidate the effect of IL-17A on microglial function. First, the cytotoxicity of various concentrations of IL-17A (10–100 ng/ml) towards BV2 cells was determined by Cell Counting Kit-8 (CCK8) assay. Compared with the controls, when the concentration of IL-17A reached 100 ng/ml, the activity of BV2 cells markedly decreased (Fig. 5A), which indicated that high-dose recombinant IL-17A in mice can reduce the activity of BV2 cells. Thus, a high level of IL-17A in the brain tissue may reduce the biological function of microglia and accelerate their apoptosis. Given that activated microglia proliferate, accumulate around Aβ plaques, and respond to neuroinflammation via phagocytosis, we further analysed the morphological changes of activated BV2 cells using a phagocytosis assay. Fluorescence analysis showed that compared with BV2 cells that were treated with Aβ alone, the number of latex beads phagocytosed by BV2 cells that were treated with IL-17A was decreased (Fig. 5B), which indicates that IL-17A weakened microglial phagocytosis. These results suggest that IL-17A may accelerate the deposition of Aβ in the brain tissues of AD model mice or patients by decreasing the phagocytosis of microglia.
Fig. 5
Effect of IL-17A on the activity and phagocytosis of BV2 cells. A Effect of IL-17A concentration on BV2 cell activity as shown by the CCK8 method. B Phagocytosis efficiency of BV2 cells incubated with IL-17A as determined with a fluorescent phagocytosis assay with latex beads. C Quantitative analysis of the number of latex beads that were phagocytosed by BV2 cells that were treated with Aβ and/or IL-17A. The values are expressed as the mean ± standard deviation (****P < 0.0001, ***P < 0.001, **P < 0.01, and *P < 0.05). The data are representative of at least three independent experiments
IL-17A promoted Aβ deposition in HT22 cellsIL-17A stimulates microglia to secrete the proinflammatory factor TNF-α, which exacerbates neuroinflammation. Another important pathological change in AD results from the deposition of Aβ plaques. To examine the effect of IL-17A on APP metabolism, we transfected HT22 cells with the mouse APP plasmid. The expression of the APP protein was markedly upregulated in APP-transfected cells compared with the control, which indicates that the recombinant plasmid was successfully constructed (Fig. 6A). Compared with the control, LPS alone, and IL-17A alone groups, the levels of Aβ1-42 in the HT22-APP-transfected cell culture medium of the LPS and IL-17A coincubated groups was significantly increased (Fig. 6B). However, no marked change was observed in Aβ1-40 (Fig. 6C). A substantial increase in the level of Aβ1-42 was observed in HT22-APP-transfected cells that were coincubated with LPS, IL-17A, and BV2 cells compared with those that were coincubated with LPS and IL-17A, but no significant change was observed in the Aβ1-40 levels in the cell culture medium (Fig. 6D). These results suggest that IL-17A and microglia strongly accelerate Aβ deposition in AD model cells.
Fig. 6
IL-17A promoted Aβ generation in HT22 cells. A Expression of APP proteins in HT22 cells that were transfected with APP overexpression plasmid and subsequently incubated with LPS and/or IL-17A for 24 h. B Aβ1-42 levels in the culture medium of HT22 cells that were transfected with APP-overexpressing plasmids and subsequently incubated with LPS and/or IL-17A for 24 h. C Aβ1-40 levels in the culture medium of HT22 cells that were transfected with APP-overexpressing plasmids and subsequently incubated with LPS and/or IL-17A for 24 h. D Aβ1-42 and Aβ1-40 levels in the culture medium of HT22 cells that were transfected with APP-overexpressing plasmids and subsequently incubated with LPS and IL-17A or BV2 for 24 h. The values are expressed as the mean ± standard deviation (****P < 0.0001, ***P < 0.001, **P < 0.01, and *P < 0.05). The data are representative of at least three independent experiments
留言 (0)