
記住我
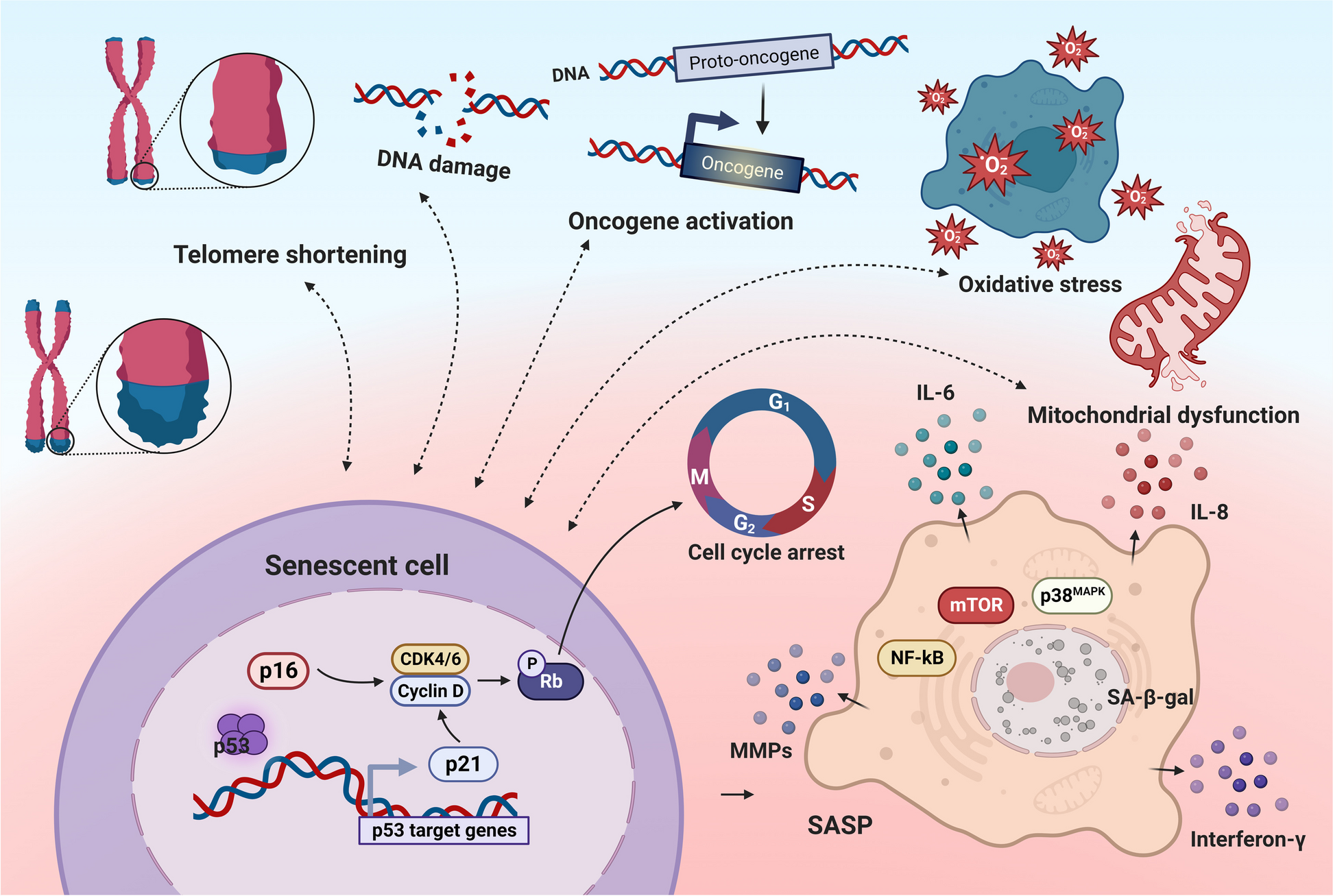
Age-related increases in senescent cells were previously believed to be associated with increased susceptibility to tumors, infectious diseases, and poor responses to vaccination [16]. In fact, middle-aged people are thought to be more likely to experience chronic fatigue under multiple stresses, which can easily induce inflammatory states and abnormal immune responses to antigens [17]. We analyzed clinical samples from Sun Yat-sen University First Affiliated Hospital over the past six years and found that the proportion of lung cancer patients increased significantly in the middle-aged age group compared to the young and elderly groups (Table 1) (Supplementary Fig. 1a). The distribution of senescent T cells in the middle-aged population is still unclear.
Table 1 Lung cancer patient samples from The First Affiliated Hospital of Sun Yat-sen University over the past six years (n = 4498)To determine the difference in the number of senescent T cells in the middle-aged group compared to the young and elderly groups, we recruited 88 healthy donors, PBMCs from the 88 donors between the ages of 26 and 86 years, we divided our donors into three groups, young group (< 45 years old,mean age = 34.1), middle-age group (45–65 years old,mean age = 54.13) and elderly group (> 65 years old,mean age = 75.57) [18]. All donors were HIV negative and free of cancer and other infection diseases. Mononuclear lymphocytes were shown by representative flow cytometry analysis of CD3 + CD8 + /CD4 + CD45RA + CCR7-CD27-CD28-CD57 + KLRG1 + expression indicating frequency of senescent T cells (Fig. 1a); The frequency of CD8 + senescent T cells (24.26 ± 3.563, N = 34) was significant increase in middle-aged group compare to the young group (10.36 ± 1.186, N = 26) and elderly group( 13.64 ± 1.177, N = 28) (p = 0.017, p = 0.0115) (Fig. 1b), similar results were observed in CD4 + senescent T cell subsets (p = 0.0329,Fig. 1c) ( young:1.075 ± 0.2275 N = 26; middle:1.564 ± 0.3059 N = 34;elderly: 0.7812 ± 0.1327 N = 28) and the proportion of CD8 + senescent T cells was higher than that of CD4 + T cell at each age group (p < 0.001, Supplementary Fig. 1b). Senescent T cell originated from effector T cells (Teff) which were at the end stage of T cell differentiation. We observed that middle-aged people had the most abundant effector T cells in CD8 + T cell population (Supplementary Fig. 1c). Additionally, we found that the frequency of CD8 + Teff cells was higher than that of CD4 + Teff cells at different age group (p < 0.05, p < 0.001, Supplementary Fig. 1d).
Fig. 1
The highest frequency of CD8 + senescent T cells in middle-age population was analyzed by FACS. a Representative flow cytometric analyses of senescent T cells (CD3 + CD45RA + CCR7-CD27-CD28-CD57 + KLRG1 +) in CD8 and CD4 T cells (top quadrant) and representative flow cytometric analyses of CD27-CD28-CD57 + KLRG1 + subsets, gated on CD3 + CD4 + /CD8 + CD45RO + CCR7-(Tem) population (bottom quadrant). The frequency of CD8 + senescent T cells (b) and CD4 + senescent T cells (c) at young group, middle-age group and elderly group. The percentage of CD57 + KLRG1 + expression on CD3 + CD8 + CD45RO + CCR7-CD27-CD28-population (d) and CD3 + CD4 + CD45RO + CCR7-CD27-CD28-population (e) at different age groups. Expressed as the mean ± SEM. *P < 0.05, **P < 0.01; Mann– Whitney test (two-tailed) and nonpaired Student’s t-test. Tem, effector memory T cell
As the thymus gradually shrinks with age, the number of naïve T cells decrease while memory T cells increase [19]. Based on the differentiation pattern of T cells in peripheral lymphoid organs as against the antigen, memory T cells differentiated into effector T cells [20]. The composition of CD27-CD28-CD57 + KLRG1 + subsets in CD3 + CD45RO + /CD45RA-CCR7- population was analyzed to determine whether the precursors of senescent T cells are present in effector memory T cell (Tem) cells (Fig. 1a). It was found that although the proportion of Tem cells was highest in the elderly group (Supplementary Fig. 1c), the frequency of CD57 + KLRG1 + double positive cells from CD8 + CD45RO + /CD45RA-CCR7-CD27-CD28- was still the highest in the middle-aged group (young: 12.66 ± 1.439 N = 26; middle: 21.74 ± 3.794 N = 34; elderly: 11.15 ± 1.422 N = 28) (p = 0.0494, p = 0.0187, Fig. 1d). The proportion of CD4 + CD45RO + /CD45RA-CCR7-CD27-CD28-CD57 + KLRG1 + T cells was roughly the same across the different age groups (Fig. 1e). Additionally, more CD57 + KLRG1 + subsets were observed in CD8 + T cells than in CD4 + T cells within the Tem subsets (Fig. 1d, e). However, the number of CD4 + Tem was higher than that of CD8 + Tem across age groups (p < 0.05, p < 0.001, Supplementary Fig. 1e).
These results indicate that the frequency of senescent T cells was highest in the middle-aged group, and the proportion of CD8 + senescent T cells was higher than that of CD4 + senescent T cells. Furthermore, the proportion of Tem cells with senescent T cell phenotype was highest in the middle-aged group.
The number of senescent T cells significantly reduced with Metformin treatmentIt is important to reduce senescent T cells through interventions to prevent the occurrence of age-related diseases, especially tumors. Metformin not only has a hypoglycemic effect but also plays a role in anti-tumor and anti-aging [21, 22]. There is little evidence supporting the modulation of T cell senescence by metformin. To investigate the effect of metformin on senescent T cells, PBMCs derived from a middle-aged population were treated with different concentrations of metformin in vitro: 0 mM, 5 mM, 10 mM, and 20 mM. The results showed that treatment with 5 mM metformin had no significant effect on senescent T cells (Supplementary Fig. 2a). The number of senescent T cells significantly decreased after treatment with 10 mM metformin (from 47.29% to 34.3%, with a 27.5% inhibition rate) (p < 0.001, Supplementary Fig. 2a). The frequency of CD8 + senescent T cells also decreased with 20 mM metformin treatment (from 36.3% to 22%, with a 39.4% inhibition rate) (p = 0.004, Fig. 2a, b). Therefore, a metformin concentration of 20 mM was determined for the subsequent research. Similar inhibition was observed in subsets of CD8 + CD45RO + CCR7-CD27-CD28-CD57 + KLRG1 + cells from Tem cells (p = 0.0005, Fig. 2a, c, Supplementary Fig. 2b). Our findings show that metformin could reduce the frequency of senescent T cells in middle-aged populations.
Fig. 2
The number of CD8 + senescent T cells in middle-age group decreased significantly after metformin treatment. (a) Frequency of CD3 + CD8 + CD45RA + /CD45RO + CCR7-CD27-CD28- cells which gated on CD3 + CD8 + T cells treated with the different concentrations of metformin (0 mM, 5 mM, 10 mM or 20 mM). Quantification of the percentage of CD8 + senescent T cells derived from Teff cells (CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57 + KLRG1 +) (b) and Tem cells (CD8 + CDR45RO + CCR7-CD27-CD28-CD57 + KLRG1 +) (c) at middle-age group with 20 mM metformin treatment. Expressed as the mean ± SEM. ***P < 0.001; Mann–Whitney test (two-tailed) and paired Student’s t-test. Teff, effector T cells; Tem, effector memory T cells; Met(-), control; Met( +), 20 mM metformin treatment
Metformin reduces the secretion of IFN-γ in CD8 + senescent T cellsSASP (senescence-associated secretory phenotype, SASP) secreted by senescent T cells could be harmful to neighboring healthy cells and even induce senescence [7]. IFN-γ is highly expressed in senescent T cells [23]. To investigate whether metformin inhibits the production of IFN-γ, PBMCs from the middle-aged group were treated with 20 mM metformin for 24 h. There was no significant difference in the production of IFN-γ from CD8 + senescent T cells between the metformin treatment and control groups (Fig. 3a, b); however, compared with the control group, the average fluorescence intensity (MFI) of IFN-γ in CD8 + senescent T cells in the treatment group was significantly decreased (P < 0.01, Fig. 3c). Moreover, the frequency of CD3 + CD8 + CD45RA + CCR7-CD27- CD28- CD57 + KLRG1 + IFN-γ + T cells in lymphocytes decreased after metformin treatment (p = 0.064, Fig. 3d). For non-senescent T cells (CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57-KLRG1-), the production of IFN-γ increased with metformin treatment (P < 0.001, Fig. 3e, f). Similar results were observed in CD4 + T cells (Supplementary Fig. 3a, b).
Fig. 3
Metformin reduces the secretion of IFN-γ in CD8 + senescent T cells. Representative flow histogram of IFN-γ production by CD8 + senescent T cells (CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57 + KLRG1 +) (a) and non- senescent T cells (CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57-KLRG1-) (e) in the control group and the 20 mM Met treatment group relative to the unstimulated controls from middle-age subjects. Analysis of IFN-γ-secreting CD8 + senescent T cells (b) and MFI of IFN-γ in CD8 + senescent T cells (c) with 20 mM metformin treatment. d Frequency of CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57 + KLRG1 + IFN-γ + T cells in lymphocytes with Met treatment and control. f Percentage and MFI of IFN-γ in CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57-KLRG1- T cells with Met treatment and control. g Representative flow histogram of IFN-γ secretion by CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57 + /CD57- population relative to the unstimulated controls from middle-age donors. h Percentage and MFI of IFN-γ in CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57 + / CD57- population. Expressed as the mean ± SEM. **P < 0.01, ***P < 0.001; Paired t test. Met, metformin
Studies have revealed that capacity of proliferation on CD57 + T cells was seriously impaired, and CD57 is considered as the best marker of cellular senescence [24]. We analyzed the differences between CD57- and CD57 + T cell subsets. More IFN-γ production in CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57 + T cells than in CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57-T cells was observed (P < 0.001, Fig. 3g, h).
Together, these results indicate that metformin inhibits the secretion of IFN-γ in senescent T cells but does not down-regulate IFN-γ production in non-senescent T cells.
Metformin inhibit proinflammatory cytokine IL-6 productionIL-6 is one of the SASP factors that are present in various types of senescent cells during aging [25]. This cytokine is primarily secreted by innate immune cells, such as monocytes and dendritic cells (DC) [26]. To identify whether metformin inhibit IL-6 production, we analyzed the secretion of IL-6 in CD3-negative cells from the peripheral blood of middle-aged subjects. The results showed that the percentage of IL-6 producers in the control group was 3.8% in CD3-cells, whereas it was only 1% in CD3-cells with metformin treatment. Additionally, the levels of IL-6 expression (MFI) in CD3- cells were significantly decreased upon metformin treatment (P < 0.001, Fig. 4a). We also compared the frequency of IL-6-producing CD3-CD57- T cells with that of CD3-CD57 + T cells, and found that the former had a higher frequency (P < 0.001, Fig. 4b). CD57, recognized as a surface marker of replicative senescence in T cells, is also expressed on maturing NK cells [27]. However, it remains unclear whether CD3-CD57 + cells exhibit a cellular senescent as well as CD3 + CD57 + T cells. we examined that the frequencies of IL-6-producing cells were reduced in both CD3-CD57 + cells (P < 0.001, Fig. 4c) and CD3-CD57- cells with metformin treatment compared to the control group (P < 0.001, Fig. 4d). Similarly, a small proportion of IL-6-producing cells was observed in CD3 + CD8 + /CD4 + T cells, and metformin treatment led to a decrease in the levels of IL-6 production. This consistent decrease was also observed at the MFI levels of IL-6 (P < 0.05, Supplementary Fig. 4a, b).
Fig. 4
Metformin inhibited the production of proinflammatory cytokine IL-6 in CD3 negative cells. Representative flow analysis of IL-6 production and MFI level in CD3- cells (a), CD3-CD57 + cells (c) and CD3-CD57- cells (d) between 20 mM Met treatment and control for 24 h in middle age group. Representative flow analysis of IL-6 production and MFI in CD3-CD57 + and CD3-CD57- population (b). Expressed as the mean ± SEM. ***P < 0.001; Paired t test. Met, metformin
Taken together, our functional analysis suggests that CD3-CD57- cells have a greater capacity to produce IL-6 than CD3-CD57 + cells, and that metformin reduces IL-6 production in both CD3- cells and T cells.
Metformin has little impact on Granzyme B secretion in senescent T cellsCytotoxic T cells (CTLs) and Natural Killer cells (NK) are responsible for producing Granzyme B (GB), which mediates cytotoxicity and kills target cells, senescent T cells express high levels of GB [4]. In this study, we investigated the effect of metformin treatment on GB production. Our findings indicate that metformin treatment leads to an increase in GB production (P < 0.05, Fig. 5a, b), while there was no significant difference of MFI (Fig. 5b) and the expression of GB in CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57 + KLRG1 + T cells from lymphocytes between the metformin treatment and control groups (Fig. 5c). In non-senescent T cells, no significant differences in GB production were observed with metformin treatment (Fig. 5d, e), although there is an increasing tendency of frequency of CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57-KLRG1-GB + T cells in lymphocytes compared to the control group (Fig. 5f). Interestingly, our results demonstrate that CD3 + CD57 + T cells, when compared to CD3 + CD57-T cell populations, secrete higher levels of GB (Fig. 5g, h). Overall, our findings suggest that metformin has little impact on Granzyme B secretion in senescent T cells.
Fig. 5
The effect of Metformin on GB secretion. Representative flow histogram of GB secretion by CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57 + KLRG1 + (a) and CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57-KLRG1- (d) in the control group and the 20 mM Met treatment group relative to the unstimulated controls group from middle-age donors. Analysis of GB production in CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57 + KLRG1 + (b) / CD57-KLRG1- (e) T cells, cultured with 0 mM or 20 mM metformin for 24 h in the middle age group. The proportion of CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57 + KLRG1 + GB + (c) and CD57-KLRG1-GB + (f) T cells in lymphocytes with metformin treatment compare to control group. g Representative flow histogram of GB secretion by CD3 + CD57 + and CD3 + CD57-T cells relative to the unstimulated controls group in middle-age donors. h Percentage and MFI of GB in CD3 + CD57 + and CD3 + CD57-T cells. Expressed as the mean ± SEM. *P < 0.05; Paired t test. Met, metformin
Metformin promotes the production of TNF-α in senescence T cellsTNF-α, tumor necrosis factor α, is an effective pro-inflammatory cytokine that plays a crucial role in the maintenance and homeostasis of the immune system, inflammation, and host defense. High concentrations of TNF-α can contribute to various pathological processes, including inflammaging, autoimmune diseases, and tumors. It has been observed that senescent T cells also secrete high levels of TNF-α [4, 28, 29]. After intervention with Metformin, an increased tendency in TNF-α secretion from CD8 + senescent T cells was observed when compared to the control group (Fig. 6a). The frequency of CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57 + KLRG1 + TNF-α + T cells in lymphocytes also significantly increased (p < 0.01, Fig. 6b). Similar results were observed in CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57-KLRG1-T cells (p < 0.01, Fig. 6c). As expected, CD8 + CD45RA + CCR7-CD27-CD28-CD57 + T cell subsets secreted significantly more TNF-α compared to the CD57- subgroup (p < 0.01, Fig. 6d). Additionally, CD4 + senescent T cells showed an increased tendency in TNF-α production with metformin treatment (Supplementary Fig. 5a). However, there was no difference in TNF-α production in CD3 + CD4 + CD45RA + CCR7-CD27-CD28-CD57-KLRG1- T cells with metformin treatment (supplementary Fig. 5b); instead, the frequency of CD3 + CD4 + CD45RA + CCR7-CD27-CD28-CD57-KLRG1-TNF-α + T cells in lymphocytes significantly decreased (p < 0.05, Supplementary Fig. 5c). Overall, our results demonstrate that metformin promotes the production of TNF-α in senescent T cells.
Fig. 6
Metformin promotes the production of TNF-α in senescence T cells. Analysis of TNF-α-secreting cells from CD8 + senescent T cells (CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57 + KLRG1 +) (a) and CD3 + CD8 + CD45RA + CCR7-CD27-CD28-CD57-KLRG1-T cells (c) in the control group and the 20 mM Met treatment group in middle-age group. b The frequency of TNF-α + CD8 + senescent T cells in lymphocytes between the control and Met treatment group. d Representative flow histogram of TNF-α in CD8 + CD45RA + CCR7-CD27-CD28-CD57 + /CD57-T cell and the percentage and MFI of TNF-α in CD8 + CD45RA + CCR7-CD27-CD28-CD57 + /CD57-T cell. Expressed as the mean ± SEM. **P < 0.01; Paired t test. Met, metformin
Metformin increased the telomerase concentration and the frequency of undifferentiated T cellsTelomere, a DNA–protein complex located at the end of chromosomes, plays a crucial role in maintaining chromosome stability. However, during cell divisions, the length of telomeres naturally decreases, ultimately leading to cell aging, death, or cancer [8]. Telomerase, an enzyme with reverse transcription activity, is responsible for synthesizing telomeric DNA and preserving the length of telomeres [8]. As T cells age, telomeres shorten and telomerase activity decreases, resulting in the loss of their proliferative ability [8]. Our above results indicate that metformin is able to reduce the number of senescent T cells, whether it has an effect on telomerase is still unclear. We collected the supernatant of PBMCs that were cultured with 20 mM metformin for 24 h in the middle-aged group and measured telomerase concentration using an enzyme-linked immunosorbent assay (ELISA). The results showed that the telomerase concentration in the control group was 0.71 ng/ml (0.71 ± 0.07, N = 28). However, with metformin treatment, the concentration of telomerase increased to 1.33 ng/ml (1.33 ± 0.24 N = 28) (P < 0.05, Fig. 7a). Immunosenescence is characterized by an increase in aged T cells and a decrease in naive T cells which possess stemness [10]. Consistent with previous studies, our results also revealed a lower proportion of CD8 + Tn (CD3 + CD8 + CD45RA + CCR7 +) cells in the elderly group (3%) compared to the middle-aged group (6%) and the young group (9%) (P < 0.05, Supplementary Fig. 6). Surprisingly, metformin increased the frequency of undifferentiated T cells (CD3 + CD4 + /CD8 + CD45RA + CCR7 + CD27 + CD28 +) (P < 0.05, Fig. 7b, c). These findings suggest that the increased telomerase concentration and number of undifferentiated T cells induced by metformin may contribute to its anti-aging effects.
Fig. 7
Metformin increased the telomerase concentration and the undifferentiated T cells. a The concentration of telomerase in medium (ng/ml) was analyzed in PBMCs with 20 mM Metformin treatment and control at middle age group, examined by ELISA. The flow cytometric analysis the frequency of CD3 + CD4 + CD45RA + CCR7 + CD27 + CD28 + (b) and CD3 + CD8 + CD45RA + CCR7 + CD27 + CD28 + (c) T cells in lymphocytes between the metformin treatment and control. Expressed as the mean ± SEM. *P < 0.05, **P < 0.01; Mann–Whitney test (two-tailed) and paired Student’s t-test. ELISA, Enzyme-linked immunosorbent assay. Met (-), control; Met ( +),20 mM metformin treatment
Metformin promoted the expression of genes related to stemness and telomerase activity, inhibited the expression of β-galactosidase (SA-β-Gal) and DNA damage associated genesTo provide a detailed characterization of cellular senescence, including senescence-associated β-galactosidase (SA-β-Gal) production, telomere shortening, decreased telomerase activity, and increased DNA damage, cell cycle arrest [12, 30], we conducted RNA-sequencing on PBMCs obtained from three middle-aged subjects. The cells were treated with metformin or served as control groups to examine the effects.
There were 26,089 differentially expressed genes (DEGs) co-expressed between the group treated with metformin and the control group; additionally, there were 2,694 specifically expressed DEGs in the metformin treatment group and 3,844 in the control group (Fig. 8a). To characterize the transcriptomic changes between metformin treatment and the control group, DEGs were displayed using a volcano plot (Fig. 8b) and expression heatmap (Fig. 8c). The gene GLB1, which is associated with SA-β-Gal, was significantly decreased in the metformin-treated group compared with the control group (Fig. 8b, c). DNA damage-associated genes, including CDKN1A, CHEK2, E2F2, NBN, and GADD45G [31,32,33,34], were downregulated with metformin treatment (Fig. 8c). In contrast, metformin promoted the expression of cyclin-related genes CCNG1 and CCND2 (Fig. 8b, c). Consistent with ELISA data from telomerase studies, the expression of ATM, ATR [35], PRKCQ [36], and MYC [10] mRNA was significantly elevated in the metformin treatment group (Fig. 8c). DEGs revealed increased expression of TCF7, SELL, CD28, CD27 [37], genes associated with stemness (Fig. 8b-d), which is consistent with flow cytometry analysis where the frequencies of undifferentiated T cells were significantly increased in the metformin treatment group (Fig. 7b, c). A detailed analysis of the DEGs was performed by the functional annotation of Gene Ontology (GO), which showed enrichment for a highly diverse array of biological processes consistent with the change of cytokine secretion with metformin treatment. Biological processes related to lymphocyte proliferation were upregulated, such as interleukin-2 (IL-2) and interleukin-4 (IL-4); however, SASP-associated biological processes, such as IL-6, IL-8, IL-1, TNF, and IFN-γ production, were decreased with metformin treatment (Fig. 8e). These results supported that metformin promoted the expression of genes related to stemness and telomerase activity, but inhibited the expression of cellular senescence-related genes.
Fig. 8
The RNA -seq analysis of gene expression associated with T cell senescence with metformin treatment. a Venn diagram portraying the intersections and disjunctive unions of differentially expressed genes in metformin treatment group (green) and control group (gray). b The Volcano Plot showed the overall distribution of genes with up-regulated expression (green), down-regulated expression (dark gray) and no significant difference (light gray) in the metformin treatment group compared to the control group. c Expression heatmap of a selection of senescence- and stemness-associated genes in PBMCs from metformin treatment group and control group at middle-age subjects (n = 3). Data are represented as Z-scores. d Representative genome browser visualizations of normalized reads at indicated gene loci in PBMCs from a representative donor before and after metformin treatment. e The chart portrayed a selection of biological process, from GO functional enrichment analysis, which were up-regulated and down-regulated with metformin treatment. All the above RNA sequencing datas came from three independent donors and three independent experiments
留言 (0)