
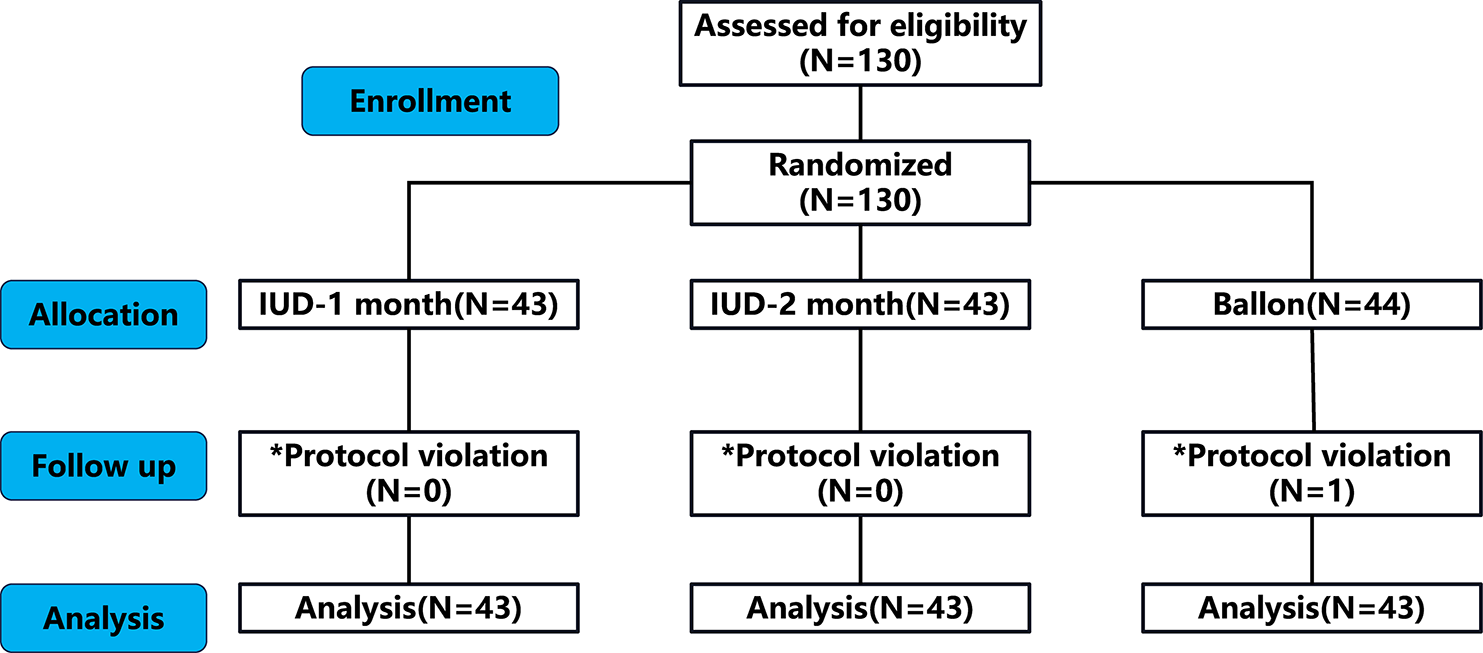
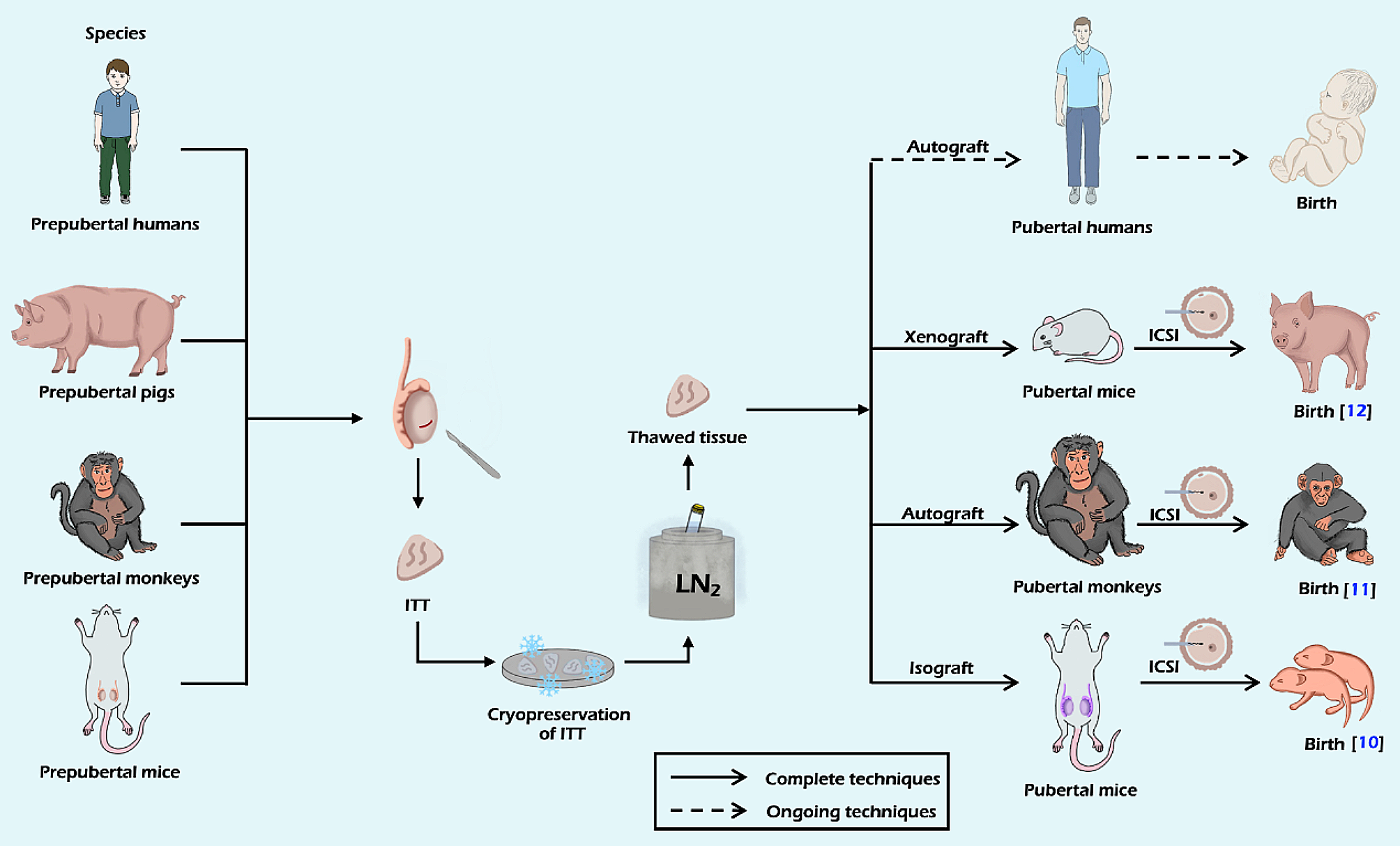
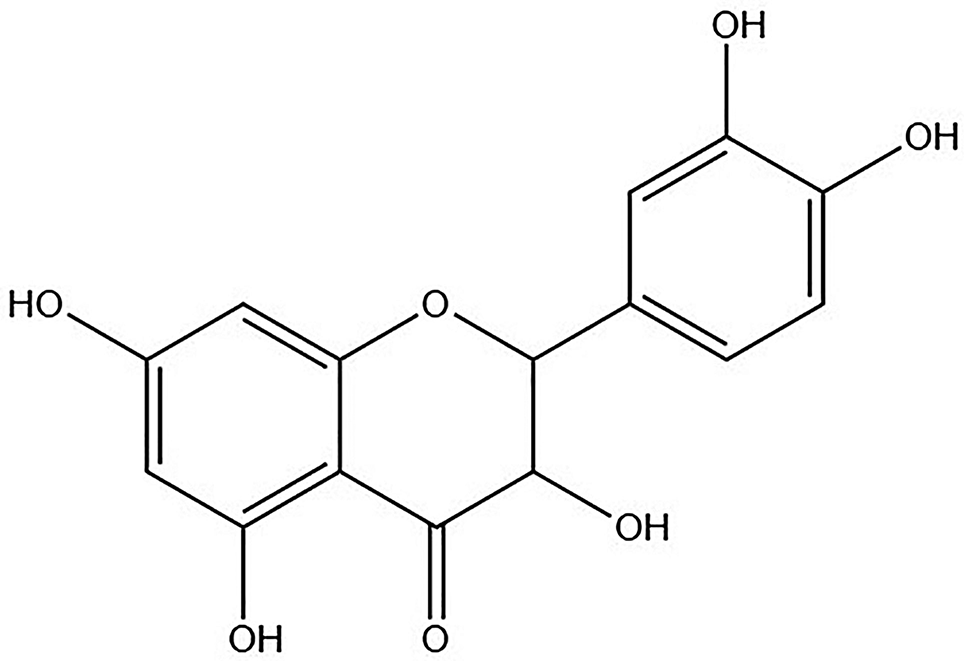
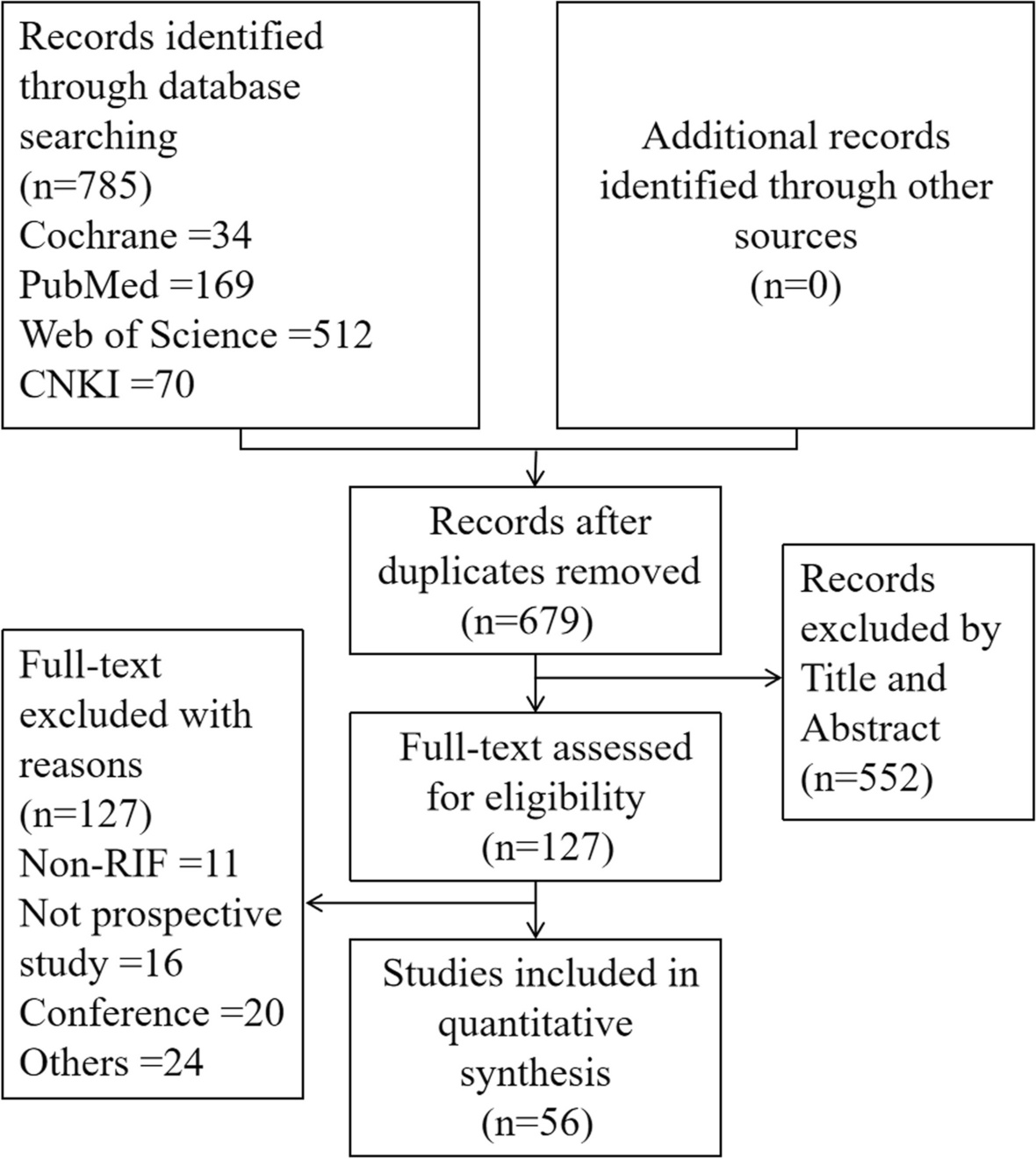
記住我

Our strategy for generating the Padi2/Padi4 DKO mouse line is shown in Fig. 1a. Padi2-null males and females were bred to generate Padi2-null zygotes and these embryos were then injected with a Padi4 sgRNA construct and transferred to wild type (FVB/NJ) recipient females to generate founder pups. The Padi4 sgRNA construct was designed to target Exon 1 of the Padi4 gene (Fig. 1b). In total, 39 founders were born, with 13 CRISPR-edited founders being identified by heteroduplex PCR, subcloning, and Sanger sequencing. All of the edited founders were heterozygous mutants, with seven of these founders having a frame-shift mutation. Following back breeding to P2KO, crossing the heterozygous F1 offspring, and genotyping successive generations, one homozygous DKO strain was established and selected for further characterization. This mutant mouse line contained a four-nucleotide deletion in Exon 1 of Padi4 (Fig. 1c) and a predicted premature stop codon (Fig. 1d).
Validation of the DKO mutant line is shown in Fig. 2. We first confirmed that P2KO and DKO mice lacked Padi2 transcripts by RT-PCR analysis of salivary gland (Fig. 2a). We next tested whether the Padi4 sgRNA targeted site in DKO mice contained a deleted sequence by performing PCR analysis of genomic DNA and then separating the DNA fragments on high percentage polyacrylamide gels. Results showed that the DKO amplicon displayed a reduced molecular weight when compared to the P2KO and WT amplicons (Fig. 2b), suggesting that DKO mice contained the targeted deletion, as was shown by Sanger sequencing of the Padi4 locus (Fig. 1c). As expected, no amplicon was observed in P4KO mice. The above results suggested that DKO mice lacked both Padi2 and Padi4 transcripts. We then performed Western blot analysis of WT, P2KO, P4KO and DKO spleen lysates to test if the DKO mice also lacked PAD2 and PAD4 protein. We probed the protein lysates with anti-PADI2 antibodies and found that, while the antibody was reactive with appropriately sized ~ 75 kDa bands in the WT and P4KO lysates, the antibody did not react with similarly sized bands in the P2KO and DKO mice (Fig. 2c, Additional file 1). Likewise, the anti-PADI4 antibody was reactive with ~ 75 kDa bands in the WT and P2KO lysates but was not reactive with similarly sized bands in the P4KO and DKO lysates (Fig. 2c, Additional file 1). These results indicate that the Padi2/Padi4 DKO strain does not express mature PAD2 and PAD4 proteins. Lastly, we found by RT-PCR that the transcripts of Padi1 and Padi3 continue to be expressed in our DKO mice, indicating that the observed phenotype was due to the disruption of Padi2 and Padi4 and that the other Padi loci remain intact (Fig. 2d).
Effect of Padi2/Padi4 DKO on fertility parametersIn order to begin testing for links between PADs and reproductive function, we first performed a controlled breeding trial to test whether a range of reproductive parameters were affected by DKO. Seven-week old mice were paired in 4 mating categories (WTxWT, DKOxDKO, DKOxWT and WTxDKO) and monitored daily for 63 days. Results showed that, although not statistically significant, DKOxDKO pairs tend to lag behind WTXWT pairs in the time taken to have their first litter (Fig. 3a). To identify which sex may be responsible for this reproductive delay, we bred DKO males with WT females and DKO females with WT males, and the trend towards decreased fertility remains more apparent for DKO males than for females (Fig. 3a).
Fig. 3
Breeding phenotype of Padi2/Padi4 double knockout mice. a In controlled breeding trials, DKO pairs (n=6) took longer than WT pairs (n=7) to have their first litter. When paired with WT mates, DKO males (n=12) took longer than DKO females (n=10) to have their first litter and had fewer pup-producing pairs (data trending, but not significant, one-tailed Fisher’s Exact Test, P = 0.19). b Unsexed pups from the breeding trial litters were weighed at two days post-partum, and showed that DKO pups were smaller than pups of either SKO strain, and significantly smaller than WT pups (one-tailed t-test, *P = 0.004). c Male DKO are significantly smaller than both SKO and WT mice at weaning (ANOVA, p=0.02, post-hoc one-tailed t-test with WT, **P = 0.003). d Male DKO weanlings are visibly smaller and less developed than SKO and WT. Growth curves are provided in Supplemental 2)
With respect to the offspring produced during the breeding trials in this report, we found that the sex ratio of homozygous DKO offspring was skewed (0.85 male:female) but did not deviate significantly from the expected equal ratio (?2 test, n = 243, P = 0.22), and that the average DKO litter size was the same as WT mice (two-tailed t-test, n = 4, P = 0.5). We also observed that unsexed 2-day old DKO pups from these same litters weighed significantly less than WT pups (Fig. 3b, n = 4 litters per strain, P = 0.004) and that DKO male weanlings weighed significantly less than WT (Fig. 3c-d, n = 21,45, P = 0.003; Fig. S2, Additional file 2). Taken together, results from these studies suggest that loss of both PAD2 and PAD4 suppresses fertility and affects offspring weight. Previous studies have shown that pup weight is also commonly reduced in sub-fertile mouse strains, including androgen receptor knockout (ARKO) mice [36] which is relevant to the research reported here. Given the growing links between PADs and hormone signaling, we predicted that the effect of DKO on both fertility and pup weight was due to disrupted hormone signaling in these mice.
Effect of Padi2/Padi4 DKO on pubertal onsetPubertal onset is thought to be primarily driven by increased testosterone production in males and one well-established external marker of pubertal development in rodents is preputial separation (PS) [25, 37, 38]. Therefore, as a further test of the hypothesis that hormone signaling is disrupted in DKO males, we next documented the timing of PS. Results show that DKO males took an average of 3.6 days longer than WT or SKO males to undergo PS (31.2 vs 27.6 days: ANOVA, P < 0.0001, Fig. 4a,b). Given that the timing of pubertal onset has been repeatedly, yet inconsistently, linked with obesity [39, 40], and been shown to differ between sexes within the same species/strain [41], we asked whether the delayed puberty in DKO males could be a side-effect of their smaller size at weaning. This does not appear to be the case, as DKO males are significantly heavier than WT and SKO at time of PS (ANOVA, P = 0.0002, Fig. 4c), and the mean weight of males, regardless of strain, was the same at day 27, which is the average day of PS in WT and SKO (ANOVA, P < 0.0001, Fig. 4d). Preputial separation is known to be an androgen-dependent process [38, 42], strongly linked to hormone signaling along the hypothalamic-pituitary-gonadal (HPG) axis [43, 44]. These results further support the hypothesis that hormone signaling may be altered in DKO males. Additionally, these observations suggest that PAD2 and PAD4 play an important role in androgen-driven sexual development in males.
Fig. 4
Pubertal onset is delayed in Padi2/Padi4 DKO male mice. a Preputial separation occurred later in DKO males than WT and SKO strains, with b the mean age of PS significantly higher (n=49, 36, 38, 46; ANOVA, P < 0.0001). c Smaller body size at weaning does not appear to be a factor, as DKO mice are significantly larger when they reach PS than all other strains (ANOVA, P = 0.0002), and d there is no significant difference in body weight between strains at day 27, the mean day of PS for WT and SKO mice (ANOVA, P = 0.5)
Effect of Padi2/Padi4 DKO on serum hormone levelsAs a more direct test of the hypothesis that hormone levels are altered in DKO mice, we next measured serum testosterone (T), luteinizing hormone (LH), and follicle stimulating hormone (FSH) in WT and DKO males. Results showed that levels of T, LH, and FSH differed significantly between DKO and WT at multiple timepoints (n = 5 per time point). Although highly variable, serum testosterone levels were lower in DKO males than WT males at all time-points, most significantly at day 48 (P = 0.05, Fig. 5). In contrast, FSH and LH levels were statistically similar between DKO and WT at the earlier time points, but at day 50 both hormones were significantly higher in DKO sera (P < 0.03). Although it is difficult to infer signaling responses from this data due to use of serum from different individual mice at each timepoint rather than sequential levels from the same mice across time,we were encouraged by the observation that serum T levels were consistently lower in DKO mice regardless of age, and that LH and FSH differed significantly between strains. Importantly, studies with ARKO mice also found that serum T levels are lower in mutant mice compared to WT mice [36]. Given this observation, our findings lend support to the prediction that PAD enzymes play a role in androgen signaling in vivo.
Fig. 5
Padi2/Padi4 DKO male mice show altered serum hormone levels. a Mean serum Testosterone levels were lower in DKO males when compared to WT males at all timepoints. b, c LH and FSH levels were significantly higher in 50-day old DKO mice, but significantly lower than WT in 90-day old mice (n=5, one-tailed t-test, *P = 0.05, **P = 0.02)
Effect of Padi2/Padi4 DKO on testis size and histologyTestis sizeHormone signaling within the HPG axis not only regulates pubertal onset, but also regulates body growth [41] and testis size [45, 46]. As a further test for potential associations between PADs and hormone signaling, we next investigated whether the total body weight and testis weight was altered in 90-day old adult DKO mice. Results showed that, while there was no difference in total body weight between the WT (m = 27.4 g) and DKO (m = 27.4 g, n = 5,2-tailed t-test, P = 1), both absolute testis weight (P = 0.002) and the gonadosomatic index (testis weight as a percent of body weight) (P = 0.037) were significantly lower in DKO mice (Fig. 6a,b). Yeh et al. found that both pup size and testis weight are reduced in AR knockout mice [36], and our finding that DKO testes are significantly smaller than WT testes in adults fits well with the hypothesis that PADs play a role in androgen signaling in males.
Fig. 6
Padi2/Padi4 DKO male mice have disrupted testicular development. In comparisons of 90-day old DKO to WT mice, DKO have a significantly smaller testes (n=5, one-tailed t-test, *P = 0.002) and b significantly lower gonadosomatic index (testes weight as % of body weight; n=5, one-tailed t-test, **P = 0.037). c Hematoxylin and Eosin stained sections of WT testis showing normal spermatogenic cell structures. d-f Hematoxylin and Eosin stained sections of DKO testis. Black arrows indicate: d apoptotic spermatogonia, e apoptotic spermatocytes and f atypical residual bodies (scale bars = 50 ?m)
Testis histologyThe observation that DKO testes are smaller than WT testes raised the possibility that testicular histology may also be defective in DKO males. To test this hypothesis, FFPE sections of adult testes were stained with H&E and scored for cellular morphology by a board-certified pathologist (Table S1, Additional file 3). Results of this analysis found that there were variable degrees of germ cell apoptosis in the DKO testes when compared to WT testes, with spermatogonia and spermatocytes being the most significantly affected (Fig. 6c-f). More specifically, apoptosis (mild/moderate grade) in spermatocytes (Fig. 6d) was noted in 5 out of 5 DKO mice compared to only 2 out of 5 WT samples. Apoptosis (mild grade) of spermatogonia (Fig. 6e) was noted in 4 out of 5 DKO mice testes compared to only 1 out of 4 WT mice. All five DKO testes also displayed moderate grade residual bodies (Fig. 6f) when compared with WT testes, which only displayed mild grade residual bodies. Sertoli, Leydig cells and interstitium were overall unremarkable. The increased rate of apoptosis and residual bodies did not appear to result in a decrease in sperm production with the lumen of most tubules being lined with histomorphologically unremarkable elongated spermatids and spermatozoa. Interestingly, in ARKO mice, spermatogenesis was arrested at the pachytene stage of meiosis and there was an increase in apoptotic-like bodies within the tubules [36] suggesting that androgen signaling is critical for normal spermatogenesis. Taken together, these results support the hypothesis that PAD2/4 signaling may play a role in androgen signaling-mediated spermatogenesis.
Effect of Padi2/Padi4 DKO on gene expression in the testisIn order to begin investigating whether PADs are associated with specific signaling pathways in the testis, and to more directly test the hypothesis that PAD2/4 play a role in regulating AR target gene expression, we next carried out RNA-Seq analysis of adult WT, P2KO, and DKO testes. Results show that 22 genes were upregulated and 19 genes were downregulated in P2KO samples when compared to WT samples (FDR cut-off 0.05, p-adj < 0.05). Additionally, we found that 263 genes were upregulated and 140 genes were down-regulated in the DKO samples compared to WT samples (FDR cut-off 0.05, p-adj < 0.05). Seventeen of the 22 genes that were significantly upregulated in the P2KO samples were also upregulated in the DKO samples. A comparison of upregulated genes is shown in Fig. 7a, with the complete lists of differentially expressed genes (DEGs) provided in Tables S2 andS3 (Additional files 4 and 5). Two hundred and forty-six genes that were uniquely upregulated in DKO testes were then subjected to DAVID gene ontology analysis. Of the 246 genes, 238 were identified in the mouse database and these genes were categorized into three domains: Cellular Components, Molecular Functions, and Biological Processes (Table S4, Additional file 6). Analysis of the Biological Processes domain finds that a majority of the DEGs are involved in reproductive functions (Fig. 7b), thus supporting the phenotypic traits we observed. In addition, analysis of the Molecular Function domain finds that nearly all of the DEGs appear to be involved in either nucleic acid binding or protein phosphorylation (Fig. 7c), thus further supporting the hypothesis that PADs play a role in regulating gene transcription. We next generated a hierarchical dendrogram and heatmap showing the top 100 most significantly up- and downregulated genes using summary counts over all pairwise tests of WT, P2KO, and DKO samples (FDR cut-off 0.05, p-adj < 0.05; Fig. 8; Table S5, Additional file 7). Results show that there is a good correlation in gene expression levels between replicates for each experimental group. Additionally, the gene expression profile for the DKO samples is distinctly different from that of the WT samples, while there are some overlapping expression patterns when comparing the DKO and P2KO samples. We next narrowed this pool down to 41 transcripts that showed at least a 2-fold difference between strains for further investigation of their known activities. For this analysis, we used The Jackson Laboratory’s Mouse Genome Informatics (MGI) databases [47, 48] and we reveiwed the current literature. Of these 41 transcripts, 15 were identified as lncRNAs, 20 were from protein coding genes, and 6 were undescribed or pseudogenes.
Fig. 7
RNA-seq analysis of gene expression in 4-month old WT and DKO testes. a Venn diagram of differential gene expression shows that 263 genes are significantly upregulated in DKO in comparison to WT, with only 17 genes overlapping the P2KO profile. Significance was set as FDR < 0.05 and p-adj < 0.05. Gene lists are provided in Supplementals 4 and 5. b-c Gene ontology analysis with DAVID 6.8 divides these upregulated genes into b Biological Processes with reproductive pathways heavily represented and c Molecular Functions almost entirely represented by genes involved in nucleic acid regulation
Fig. 8
Heatmap and unsupervised hierarchical dendrogram illustrating the top 100 differentially expressed genes between 4-month old WT, P2KO and DKO testes. Significance was set as FDR < 0.05 and p-adj < 0.05. Red = upregulated, Blue = downregulated. The list of DEGs used to create the heatmap is provided in Supplemental 7
lncRNAs are generally known to be involved in transcriptional regulation through either binding DNA directly or changing methylation patterns on chromatin. Alternatively lncRNAs can directly interact with other chromatin modifiers to recruit or deflect their action on transcription [49, 50]. One lncRNA worth mentioning here, Snhg12, has been linked to regulation of the cell cycle and promotion of the Epithelial-Mesenchymal Transition in embryogenesis [51]. In addition, Snhg12 has been shown to promote cell proliferation and inhibit apoptosis in multiple cancers [52], including testosterone-sensitive prostate cancer [53]. Although we do not propose a mechanism, the almost 3-fold decrease in Snhg12 levels in our DKO mice suggests loss of this factor may have played a significant role in the increased apoptosis and atypical residual bodies that are seen in Fig. 6.
The 20 protein coding genes showing more than a 2-fold change in expression between DKO and WT included multiple members of two important gene families. First, the Kallikreins are a family of serine peptidases, many of which have been linked to AR expression and progression of androgen sensitive prostate cancer cells [54]. Perhaps the best-known member of this family is prostate-specific antigen (KLK3), whose expression is mainly induced by androgen and is transcriptionally regulated by the androgen receptor [55]. Additionally, the kallikrein gene locus is highly responsive to steroid hormones, having at least 14 functional HREs in this region. In fact, many researchers use kallikreins as markers of hormone receptor activity [56]. In our DKO mice, Klk21, Klk24 and Klk27 are increased 3-fold compared to WT, and these three kallikreins have been shown to be expressed exclusively in Leydig cells and be responsive to testosterone [57]. Additionally, these 3 kallikreins are upregulated in the FOXa3 KO mouse model which shows severe testicular degeneration, increased gonadal apoptosis and decreased fertility [57]. The second over-represented group in our DKO mice are Zinc-finger proteins, a diverse family generally known as regulators of gene expression. Six Zinc-finger proteins are differentially expressed in our DKO mouse testes, two of which are of particular interest here. Zfp982 has previously been shown to be involved in cell lineage differentiation in embryogenesis [58] and the 12-fold decrease in levels of this factor in our DKO mice may have significantly disrupted developmental processes. Zfp979 (also known as Ssm1b) initiates DNA methylation and inhibits transcription in undifferentiated embryonic stem cells as well as in adult murine germ cells [59]. The 20-fold decrease of Ssm1b seen in our DKO testes could allow expression of a multitude of proteins detrimental to early development and germ cell maturation.
Within the remaining differentially expressed coding genes, several stand out as relevant to our discussion for their potential roles in steroid signaling and the HPG axis. Runx3 is decreased 6-fold in our DKO testes. This factor has been shown to regulate steroidogenesis and gonadal development in female mice [60]. Whether it acts similarly in male mice is currently unknown. Mutations in Spry4, which is upregulated 3-fold in our DKO testes, have been implicated in incomplete sexual maturation and infertility due to decreased GnRH activity in human males [61]. Sdc3 (upregulated 12-fold) is considered a regulator of obesity and has been proposed as a modulator of gonadal steroid function [62]. And finally, altered levels of Mrto4 in Balb/C mice (upregulated 3 fold in our DKO) have been linked to disruptions in spermatogenesis, fertility and testosterone levels in mice [63]. These findings further support our hypothesis that PADs play an important role in regulating gene expression in the testis, and more specifically, in regulating AR-mediated gene expression.
留言 (0)