
記住我
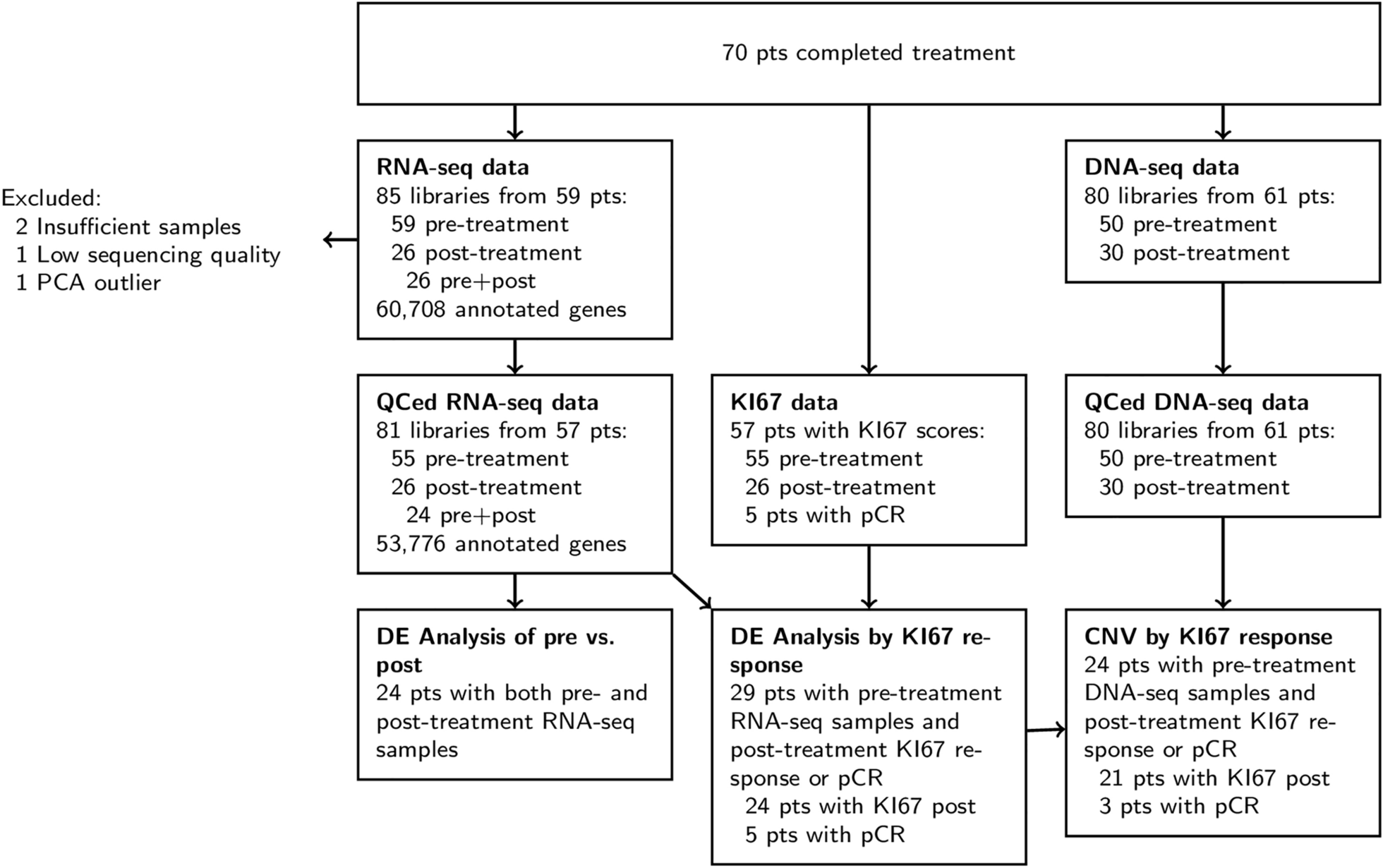




Firstly, the preliminary analysis of the expression of SMYD family members in different cancer types was carried out in Gepia online analysis website(http://gepia.cancer-pku.cn/), and the results showed that the expression of SMYD1 and SMYD4 among SMYD family was lower than control group in breast cancer tissues, whereas the expression of SMYD2, SMYD3, SMYD5 was higher than control group(Supplemetary information: Fig.S1).
Combined with the current state of research, we found that SMYD4 may have a correlation on the progression of breast cancer, but there are fewer existing studies, so SMYD4 was used as a target gene for subsequent research.
The expression of SMYD4 gene in different cancer types was further validated by the TCGA pan-cancer database(Fig. 1A). The result showed that SMYD4 expression was significantly down-regulated in TCGA and GTEx database(T = 1085,N = 291)(Fig. 1B).
Fig. 1
SMYD4 was downregulated in breast cancer. (A)The SMYD4 expression level in multiple cancer types. (B)The SMYD4 expression level in breast cancer tissues compared with normal tissues. The survival analysis of breast cancer patients with high or lowSMYD4 expression levels, including cumulative DMFS (C), OS (D), and RFS (E), predicted by the KM plotter. The SMYD4 expression level in breast cancer cell lines detected by RTqPCR (F) and western blot (G). All experiments were repeat three times (n = 3), compared variances using F test and analyzed using an unpaired t-test, error bars represent SD.*p < 0.05; **p < 0.01;***p < 0.001
Then the prognostic impact of SMYD4 in breast cancer was analyzed via the KM-plotter website, which showed that high SMYD4 expression levels were associated with better prognosis, and this correlation differed significantly in DMFS and RFS, but not in OS(Fig. 1C-E).
Finally, RT-qPCR and western blot were conducted to detect SMYD4 mRNA and protein levels in breast cell lines, including normal breast cell line MCF10A and breast cancer cell lines MDA-MB-231, CAL-51, SKBR3, MCF-7, and T47D, indicating that SMYD4 was decreased in all breast cancer cell lines as compared to MCF10A cells(Fig. 1F).
Interestingly, the expression level of SMYD4 was much lower in triple-negative breast cancer cells and CAL-51 and lumial A type cancer cells MCF-7, compared with SKBR3, MDA-MB-231 and T47D cells(Fig. 1G). In addition, the METABRIC database was used to analyse the expression of SMYD4 in different subtypes of breast cancer, but the variation between each subtype was not significant. Indicating that SMYD4 expression in breast cancer cell lines is not following with conventional Luminal type.
Taken together, these results suggest that SMYD4 is downregulated in breast cancer tissues and breast cancer cell lines, and low SMYD4 expression is associated with poor prognosis.
SMYD4 inhibits cell proliferation and invasionTo evaluate the effect of SMYD4 on breast cancer proliferation, SMYD4-FLAG overexpressed CAL-51 cells were constructed and verified by western blot(Fig. 2A).
Fig. 2
SMYD4 inhibits cell proliferation and invasion. (A) The construction of SMYD4overexpressed CAL-51 cells validated by western blot. The cell proliferation assays and invasion assays were conducted, including, colony formation assay (B), transwell assay (C), EdU assay (D), scratch assay (E) and CCK-8 (F). The apoptotic cells were analyzed through cell cytometry(G). (H)The expression levels of EMTrelated biomarkers, including Ecadherin, Ncadherin, and Vimentin in SMYD4overexpressed CAL-51 cells detected by western blot. (I) The image of tumor volume after in situ injection in SCID mice. (J) The data of tumor volume growth after in situ injection in SCID mice. (K) The protein expression level of SMYD4 in xenograft by IHC. All experiments were repeat three times (n = 3), compared variances using F test and analyzed using an unpaired t-test, error bars represent SD. **p < 0.01;***p < 0.001
SMYD4 overexpressed cells exhibited lower rates of cell clone formation, less cell proliferation rate and lower EDU positiv ratio compared to control cells, suggesting that SMYD4 could inhibit breast cancer proliferation(Fig. 2B, D,F).
Meanwhilie SMYD4 overexpressed cells showed less aggressive in transwell and scratch assays(Fig. 2C, E).
Cell flow assay shows SMYD4 promotes apoptosis in breast cancer cells(Fig. 2G).
Furthermore, western blot showed that SMYD4 overexpressed CAL-51 cells expressed more E-cadherin, less N-cadherin, and less Vimentin, while SMYD4 knockdown MDA-MB-231 cells were associated with less E-cadherin, more N- cadherin, and more Vimentin, indicating that SMYD4 is negatively associated with EMT(Fig. 2H).
To assess the role of SMYD4 in vivo, SMYD4 overexpressed CAL51 cells were injected in situ in SCID mice, which showed that SMYD4 inhibited tumor proliferation(Fig. 2I, J,K).
These cell function assays showed the same results in SMYD4 overexpressed MCF-7 cells as well as SMYD4 knockdown MDA-MB-231 cells(Supplemetary information: Fig.S2-S3).
These results suggest that SMYD4 can inhibit breast cancer proliferation and invasion both in vivo and in vitro.
SMYD4 interacts with MYH9Immunoprecipitation was performed and followed by mass spectrometry to further determine the mechanism of SMYD4 function. A total number of 349 SMYD4 binding proteins were identified, among which, MYH9 ranked at the leading and was selected as a cancer related target gene(Fig. 3A).
Fig. 3
SMYD4 interacted with MYH9. (A) The top 10 interacting genes identified by mass spectrometry. (B) The predictive protein binding model of SMYD4 and MYH9. (C) The intracellular colocalization of SMYD4 and MYH9 visualized by IF and analysis by co-localizasion ratio. The interaction of SMYD4 and MYH9 was verified by coIP assay, including endogenous (D, E) and exogenous (F). All experiments were repeat three times (n = 3)
The protein binding of SMYD4 to MYH9 was then predicted, analysed and visualised by GRAMM, PDBePISA and Pymol2 software, and the binding model of SMYD4 and MYH9 was presented as a cartoon structure(Fig. 3B).
In addition, the co-localisation levels of SMYD4 and MYH9 in breast cancer cells were analysed by immunofluorescence and fluorescence co-localisation analysis(Fig. 3C).
In addition, the interaction between SMYD4 and MYH9 was verified by endogenous and exogenous immunoprecipitation experiments(Fig. 3D-F). These results suggest that SMYD4 can directly interact with MYH9 at the protein level.
SMYD4 acts by inhibiting the wnt signalling pathwayThree groups of SMYD4 overexpressed CAL51 cells and control cells were subjected to RNAseq analysis. The analysis showed 277 up-regulated genes and 408 down-regulated genes(Fig. 4A).
Fig. 4
SMYD4 suppressed Wnt signaling by inhibiting the nuclear localization of βcatenin. (A) RNAseq differential gene volcano map between SMYD4 overexpressed and control group. The GO analysis was performed among up-regulated genes (B)and down-regulated genes(C). (D)TOP5 up- and down-regulated gene sets after GSEA analysis and the enrichment results of WNT signalling pathway(E). (F)The βcatenin expression level in the nucleus and cytoplasm detected by nucleocytoplasmic separation western blot. (G)The TOP/FOP luciferase reporter assay in SMYD4overexpressed CAL-51 cells and in SMYD4 knock-down MDAMB-231 cells. (H) The localization transformation of βcatenin visualized by IF. (I)The expression levels of downstream target genes of Wnt/βcatenin signaling, including MYH9,N-cadherin, Vimentin, C-myc, Cyclin D1 and Survivin detected by western blot. All experiments were repeat three times (n = 3), compared variances using F test and analyzed using an unpaired t-test, error bars represent SD. **p < 0.01;***p < 0.001; ****p < 0.0001
GO analysis of these differential genes showed that up-regulated genes were enriched in endoplasmic reticulum lumen, and down-regulated genes were mainly enriched in blood vessel morphogenesis an morphogenesis of an epithelium(Fig. 4B, C).
All the sequenced genes were then subjected to GSEA analysis and the enrichment results showed that the WNT signalling pathway was significantly inhibited in the SMYD4 overexpressed group(Fig. 4D, E). Therefore, the Wnt signalling pathway is considered to be a potential major pathway through which SMYD4 exerts its function.
As extensively studied, Wnt/β-catenin signalling is the classical pathway of Wnt signalling, starting with the nuclear localization of β-catenin.
Nucleoplasmic separation assay showed that in SMYD4-overexpressing CAL-51 cells, β-catenin expression was decreased in the nucleus and increased in the cytoplasm(Fig. 4F).
To assess the regulatory effect of SMYD4 on Wnt signalling, TOP/FOP luciferase assay was performed. The results showed that SMYD4 overexpression decreased TOP/FOP luciferase activity, whereas SMYD4 down-regulation increased TOP/FOP luciferase activity(Fig. 4G).
In addition, the effect of SMYD4 on the localisation of β-catenin was explored using immunofluorescence assays. The results showed that in SMYD4 overexpressed CAL-51 cells, nuclear localisation of β-catenin was significantly reduced, whereas β-catenin was abundantly present on the cell membrane(Fig. 4H).
Moreover, the expression of Wnt/β-catenin downstream target genes was detected by western blot, and it was found that SMYD4 down-regulated the expression of MYH9 protein and WNT pathway-related proteins, N-cadherin, Vimentin, C-myc, CyclinD1, and Survivin proteins(Fig. 4I).
In contrast, the expression of MYH9 and WNT pathway-related proteins was upregulated in SMYD4 knockdown MDA-MB-231 cells(Fig. 4I).
Besides, altered WNT pathway proteins due to SMYD4 overexpression were also detected in xenografts(Supplemetary information: Fig.S4).
The above results suggest that SMYD4 exerts cancer inhibition by suppressing the expression of MYH9 and inhibiting the WNT signalling pathway.
MYH9 rescues SMYD4 function in breast cancerTo further validate the role of SMYD4 in relation to MYH9, rescue experiments were performed to identify MYH9 as a key protein for SMYD4 to exert its function.
Cell clone formation assays illustrated that overexpression of MYH9 reversed the inhibition of clone formation rate, and EDU positivity in SMYD4 overexpressed cells, whereas knockdown of MYH9 rescued the facilitation funcation in SMYD4 knockdown cells(Supplemetary information: Fig.S5 A, B,F, H).
Similarly, MYH9 overexpressed cells showed function rescue in transwell and scratch assays(Fig.S5 C, E). Knockdown of MYH9 restored the increase in cell invasiveness after SMYD4 knockdown(Fig.S5 D, G).
In addition, MYH9 rescued SMYD4-induced changes in the WNT signalling pathway.
Nuclear localisation of β-catenin was restored after both MYH9 overexpression and knockdown(Fig.S5 I, J), and the addition of MYH9 also reversed the changes in TOP/FOP luciferase activity after SMYD4 overexpression and knockdown treatments(Fig.S5 K, L). The WNT pathway-related proteins N-cadherin, Vimentin, C-myc, Cyclin D1, and survivin, which were detected to be altered in the SMYD4 overexpression as well as knockdown groups, were also rescued after MYH9 overexpression and knockdown, respectively(Fig.S5 M).
SMYD4 functions through the TPR domain combined with the Ccoil domain of MYH9To further investigate the mechanism of direct interaction between SMYD4 and MYH9, truncation mutants with different domains were constructed for SMYD4 and MYH9 for subsequent studies.
Taking into account the literature and protein domain prediction websites, the 5‘TPR domain, SET domain and 3’CTD domain of SMYD4 were truncated and fused to express the HIS tag. SMYD4 full-length plasmid and three truncations of SMYD4 were co-transfected with MYH9-HA into 293T tool cells for CO-IP assays.
By CO-IP assay, SMYD4 full-length, delSET and delCTD could immunoprecipitate the MYH9-HA protein, however delTPR could not. It suggests that the 5’TPR domain of SMYD4 the domain directly interacts with the MYH9 protein(Fig. 5A).
Fig. 5
SMYD4 functions through the TPR domain combined with the Ccoil domain of MYH9. (A)Diagrammatic representation of SMYD4 and its truncated forms. 293T cells were transfected with the indicated constructs subjected to immunoprecipitation with anti-HA (against MYH9). The red boxes represents the pull-down bands. (B) Diagrammatic representation of MYH9 and its truncated forms. 293T cells were transfected with the indicated constructs subjected to immunoprecipitation with anti-HIS (against SMYD4). The red boxes represents the pull-down bands. SMYD4 delTPR rescues SMYD4-inhibited cell proliferation and invasion including colony formation assay (C), EdU assay (D), transwell assay (E), scratch assay (F). (G)The expression levels of downstream target genes of Wnt/βcatenin signaling, including MYH9, N-cadherin, Vimentin, C-myc, Cyclin D1 and Survivin detected by western blot. All experiments were repeat three times (n = 3), compared variances using F test and analyzed using an unpaired t-test, error bars represent SD.*p < 0.05; **p < 0.01; ***p < 0.001
After that, MYH9 was designed as SH3-like, delCoil, and IQCcoil truncations and fused to express HA tags, MYH9 full-length plasmid and three truncations were co-transfected with SMYD4-HIS plasmid into 293T tool cells for CO-IP, and the results showed that only IQCcoil truncation successfully immunoprecipitated the SMYD4-HIS protein, showing that the Ccoil structural domain of MYH9 is the direct binding site for SMYD4(Fig. 5B).
To verify that the TPR domain of SMYD4 is the core structural domain exerts protein function, we designed rescue experiments on the truncated protein of TPR. SMYD4 full-length and SMYD4 delTPR were transfected into breast cancer cells to verify the function.
Consistent with the hypothesised results, truncated TPR SMYD4 was unable to exert an inhibitory effect on the rate of clone formation, cell proliferation and EDU positivity, as well as an impediment to invasiveness in transwell and scratch assays(Fig. 5C-F).
Similarly, SMYD4 delTPR no longer inhibited MYH9 expression as well as the expression of WNT pathway-related proteins(Fig.S5 G). In addition, SMYD4 delTPR also rescued the effects on β-catenin nuclear localisation and the inhibitory function of TOP/FOP luciferase activity(Fig. 6A, B).
Fig. 6
SMYD4 acts by methylating MYH9 protein to promote ubiquitination degradation of MYH9. (A)The localization transformation of βcatenin visualized by IF after rescue treatment by SMYD4 delTPR (B)The TOP/FOP luciferase reporter assay after rescue treatment by SMYD4 delTPR. (C) MYH9 lysine methylation levels after SMYD4 and SMYD4delTPR overexpression detected by immunoprecipitation and western blot. (D) MYH9 lysine methylation levels after SMYD4 knockdown detected by immunoprecipitation and western blot. (E) Changes in MYH9 protein expression levels over time after CHX treatment detected by westernblot between SMYD4 overexpression and control groups. (F) After CQ and MG132 treatments, MYH9 protein expression levels in SMYD4 overexpression and control groups were detected by westernblot. Ubiquitination levels of MG132-treated MYH9 proteins detected by immunoprecipitation and western blot in SMYD4 overexpression group(G) and SMYD4 knockdown group (H).(I) Number of MYH9-bounded CTNNB promoter regions detected by CHIP and real-time qPCR. All experiments were repeat three times (n = 3), compared variances using F test and analyzed using an unpaired t-test, error bars represent SD.*p < 0.05; **p < 0.01;***p < 0.001; ****p < 0.001
The above results indicate that SMYD4 binds the Ccoil domain of MYH9 through the TPR domain and affects the expression of MYH9 protein to inhibit the WNT signalling pathway.
SMYD4 acts by methylating MYH9 protein to promote ubiquitination degradation of MYH9 and affects its binding to the CTNNB1 promoterSMYD4 is a histone and non-histone methylation transfer protein, so it is presumed that the mechanism of SMYD4 regulation of downstream MYH9 proteins is through non-histone methylation. Pan-lysine methylation antibodies purchased from Abclonal(China) were used to examine methylation of MYH9 by SMYD4.
Immunoprecipitation of MYH9 protein from CAL-51 cells transfected with full-length SMYD4 and SMYD4 delTPR was performed to detect the level of panmethylation at the location of MYH9, which showed that SMYD4 overexpression markedly increased the level of lysine mono-methylation of MYH9 but did not alter the level of lysine di-methylation and tri-methylation of MYH9, and SMYD4 delTPR did not affect any methylation level of MYH9(Fig. 6C).Similarly, the lysine monomethylation level of MYH9 was significantly reduced in the SMYD4 knockdown group(Fig. 6D).
Protein stability assays were then performed in SMYD4-overexpressed CAL51 cells, and MYH9 degradation was faster in the SMYD4-overexpressed group compared to the control group after the addition of CHX(Fig. 6E). To investigate the ubiquitination pathway through which SMYD4 promotes the degradation of MYH9, MG132 and CQ addition treatments were performed respectively, The results showed that the degradation rate of MYH9 was significantly altered by the addition of MG132(Fig. 6F), suggesting that MYH9 is degraded via the proteasome pathway.
MYH9 immunoprecipitation assay was performed on different groups of cells after 8 h of MG132-treated culture.The results showed that SMYD4 overexpression significantly increased the level of ubiquitination of MYH9 and was rescued by SMYD4 delTPR(Fig. 6G). Meanwhile knockdown of SMYD4 significantly reduced the ubiquitination level of MYH9(Fig. 6H).
In addition we verified by CHIP-qPCR that overexpression of SMYD4 significantly reduced the binding of MYH9 to the CTNNB1 promoter region, and the effect could be similarly reversed by SMYD4 delTPR(Fig. 6I).
留言 (0)