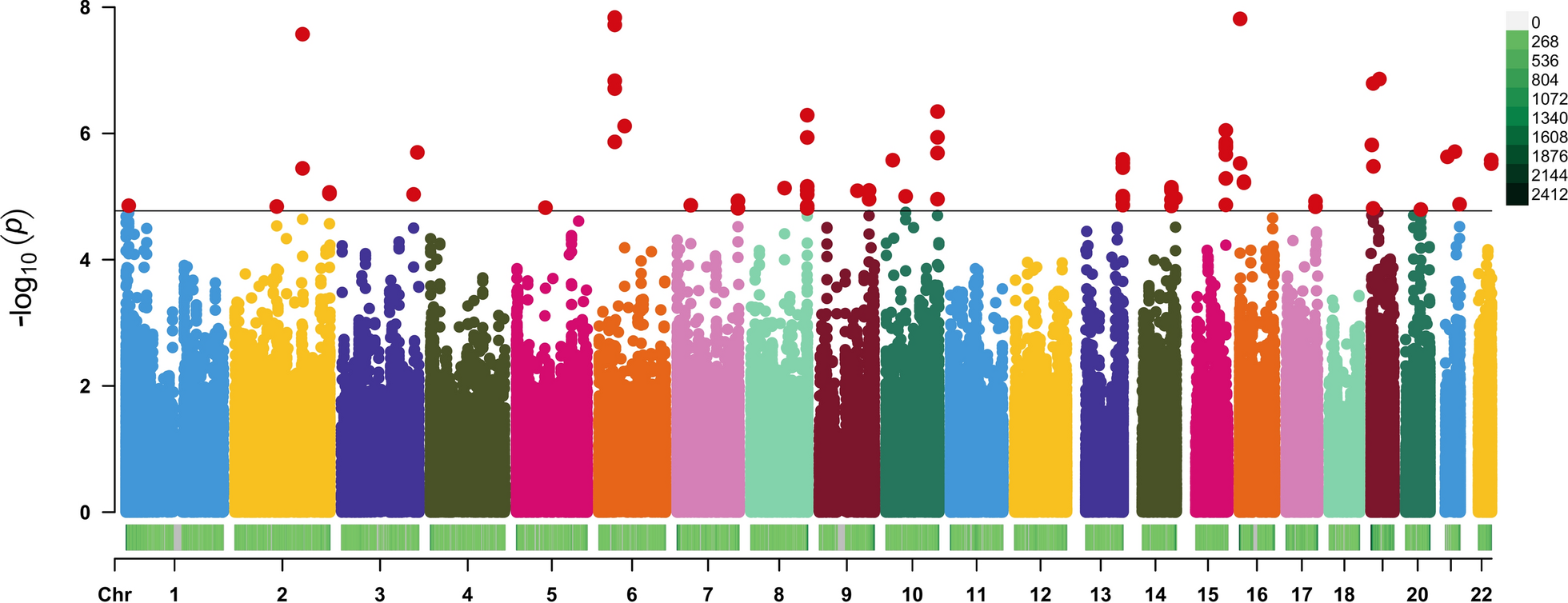
In this study, we performed a monozygotic twins-based EWAS to assess epigenetic variations associated with eGFR. We identified 80 CpGs, 34 genes, 28 DMRs, and various biological pathways associated with eGFR. In addition, we found a causal effect of DNAm on eGFR and eGFR on DNAm. The candidate gene SYNGR3 was validated in an independent community cohort. These findings contribute to further epigenetic studies of eGFR changes and provide insights for interventions and treatments for eGFR decline.
We found a negative association between the methylation level of SYNGR3 and eGFR. In an independent community population, our validation result was consistent with this. Furthermore, we demonstrated that the CpGs of SYNGR3 have a causal effect on eGFR using ICE FALCON. Similarly, a genetic study involving millions of samples also found an association between SYNGR3 and eGFR [33]. SYNGR3 was presumed to be an epigenetic regulatory gene due to its typical CpG features [34]. The SYNGR3 encoded synaptogyrins-3, a protein that regulated neurotransmitter release [35]. Synaptogyrins-3 affected renal nerve signaling by influencing the release of neurotransmitters. Renal nerves played an important role in the regulation of renal function, including glomerular filtration, sodium reabsorption, and renin release [36]. We also found a significant causal effect from eGFR on the methylation of SYNGR3. However, the exact mechanism is unclear, and further studies are needed to elucidate it.
In this study, methylation levels of CpGs located at genes such as EYS, MLLT1, and KIF26A were found to be positively associated with eGFR. Multifactorial analysis showed that high EYS expression predicted worse renal function and shorter survival in patients with clear cell renal cell carcinoma [37]. Abnormal expression of MLLT1 during early kidney development enhanced transcription, leading to the occurrence of nephroblastoma [38]. KIF26A encoded an unconventional motor protein that affected cilia formation and function, and its impairment could lead to congenital disabilities such as renal and urinary tract abnormalities [39].
We found that the methylation levels of CpGs in LONP1, NR2F2, BRD1, LITAF, and SHH were negatively correlated with eGFR. NR2F2 reduced renin promoter activity, and the consequent low renin, hypotension, and hyponatremia could lead to decreased glomerular filtration rate and renal function impairment [40]. Increased LONP1 expression has been observed in diabetic nephropathy patients and was closely associated with renal tubulointerstitial fibrosis. When LONP1 expression was inhibited, serum creatinine and renal tubule injury were improved [41]. BRD1 was found to be positively associated with urinary albumin excretion, and high urinary albumin excretion was indicative of impaired kidney function [42]. LITAF induced the secretion of tumor necrosis factor-alpha and other inflammatory mediators, causing renal inflammation and the progression of CKD[43]. SHH has been found to promote kidney damage and fibrosis [44]. In a GWAS on 1.2 million individual sample, SHH and was also found to be negatively correlated with eGFR levels, which supports our results to some extent [45]. However, the association of other genes (e.g., OLIG2, PRIMA1, LIPI, and GPR144) with eGFR has not been extensively investigated, and further studies are needed to determine their roles.
In addition, an EWAS conducted on a Korean population found that the methylation levels of CpGs located in ZNF696, GPR144, and KIF26A were associated with eGFR, consistent with our findings [46]. Another epigenome study involving 1.5 million European individuals also supported our results, identifying associations between eGFR levels and the methylation levels of CpGs in genes such as OLIG2, DLX2, ZNF493, LONP1, EBF3, ZNF696, EYS, NR2F2, SF3A2, CPN2, GRTP1, BRD1, MLLT1, LITAF, PRIMA1, GPR35, EPHB3, DRGX, KIF26A, SHH, ITGB2, CAMTA1, CD8B2, GFM2, and MAFB [33].
Gene MANIA results showed that SYNGR3 was co-expressed with MLLT1 and had gene interactions with AFF4 and EDIL3. SYNGR3 can regulate synaptic function and affect neuronal messaging. MLLT1 shift code deletion was associated with glioblastoma. AFF4 was involved in mediating the genesis and development of neurons. EDIL3 probably affected neural function through extracellular matrix accumulation. This explained possibly the interaction of SYNGR3 with MLLT1, AFF4, and EDIL3 [35, 47, 48].
Using the ICE FALCON method, this study found that eGFR was causally related to CpGs on several genes, such as C9orf3, EPHB3, and MLLT1. The aminopeptidase produced by C9orf3 was an important component of the renin-angiotensin system and caused hypertension by promoting the conversion of angiotensin II [49]. The association of hypertension with decreased eGFR and renal injury was self-evident[50]. Receptor signaling by EPHB3 regulates the cytoarchitecture and spatial organization of adult renal medullary tubular cells via Rho family GTPases and may, therefore, influence tubular reabsorption capacity [51]. In patients with nephroblastoma, MLLT1 was observed to act as a messenger to drive aberrant expression of target genes by mediating phase separation and protein–protein interactions [52].
This study identified 28 DMRs associated with eGFR within genes CDCP1, SHANK1, HOXD3, IRX1, GPIHBP1, PCDHA, PTPRG, etc. For SHANK1 and E2F8, the results of this study were consistent with previous research. The DNAm level of SHANK1 has been shown to be associated with eGFR in African HIV patients, suggesting its potential impact on kidney function [53]. The DNAm level of E2F8 has been found to be related to survival in kidney cancer patients, and E2F8 was also involved in the repair process of acute kidney injury [54, 55]. High CDCP1 expression was found to be negatively correlated with eGFR by co-expression analysis [56]. Overexpression of HOXD3 inhibited the proliferation, invasion, and migration of 786-O and CAKI-1 cells, and it was a key gene for inhibiting the progression of renal cell carcinoma [57]. IRX1, when under-expressed, promoted inflammatory responses, thereby impairing kidney function [58]. GPIHBP1 has been found to induce chronic kidney failure by impairing lipoprotein lipase function [59]. Hypermethylation of PCDHA has been demonstrated to be associated with nephroblastoma [60]. PTPRG exerted an antitumor effect by modulating the immune phenotype of renal cell carcinoma patients [61].
This study has several strengths. First, the use of monozygotic twin design controls for genetic background, family upbringing, and intrauterine environment, thereby enhancing the credibility of the study findings. Second, we conducted causal inference and identified that DNAm has a causal effect on eGFR. Thirdly, using the Chinese population as a sample, this study helps to provide information on the decline in eGFR in Chinese.
However, this study also has some limitations. First, the sample size of this study was relatively small due to the difficulty of obtaining twin samples. However, this study employed a trait-discordant twin design, significantly reducing the required sample size compared to traditional cross-sectional or case–control designs while achieving the same statistical power [62]. Additionally, although every effort has been made to control for confounding factors, some unknown factors may affect the results that cannot be measured.






留言 (0)