
記住我

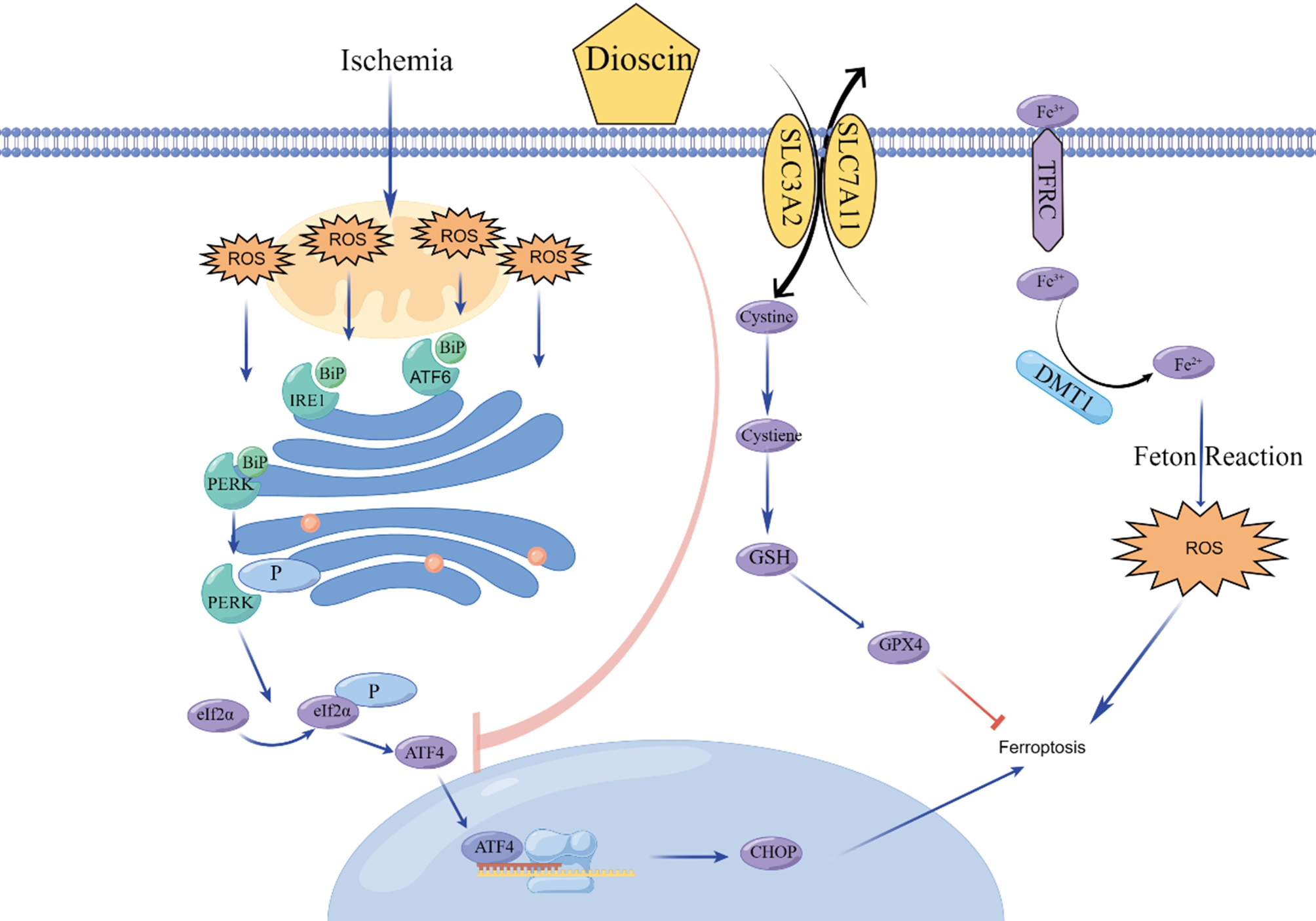



IMP could inhibit the proliferative and migratory activities of PCa. A Analysis of histidine and risk of death from PCa. B Cell viability assay of PC3 and DU145 cells following HIS stimulation. C Cell viability assay of PC3, DU145, LNCap and 22RV1 cells after IMP stimulation. D Inhibition rate of IMP (10 mM) on PC3 and DU145 cells at different times. E Cell colony formation of PC3 and DU145 cells with IMP stimulation. F EdU staining plots of PC3 cells after IMP stimulation. Scale bar, 200 μm. G Wound healing diagram after IMP stimulation of PC3 cells. Scale bar, 500 μm. H Wound healing diagram after IMP stimulation of DU145 cells. Scale bar, 500 μm. I Transwell migration plots after IMP stimulation of PC3 and DU145 cells. Scale bar, 200 μm. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001; ns, not significant
To demonstrate the relationship between His metabolic and PCa, we conducted an analysis of the correlation between His levels and PCa using a comprehensive biomarker-disease association atlas (available at nightingalehealth.com/atlas). The results showed that His concentration was negatively correlated with PCa mortality [Hazard ratio (HR) = 0.768, P = 2.48E−05] (Fig. 1A and Table S2). Additionally, we treated two commonly used CRPC cell lines (PC3 and DU145) with His for 24 h and 48 h, respectively, as a way to determine the exact relationship between His and CRPC. The results of CCK8 assay showed that the cell viability of both cell lines remained above 75% with the increment of His concentration (Fig. 1B). We therefore speculate that the negative correlation between His and PCa may be related to its metabolites.
Subsequently, we investigated whether IMP was able to influence the proliferation of PCa cells using a cell viability assay to determine whether IMP was able to influence the proliferation of PCa cells. The results showed that the viability of PC3 and DU145 cells decreased significantly with increasing IMP concentration (Fig. 1C). Nevertheless, the viability of LNCap and 22RV1 cells remained above 80% throughout the experimental period. Subsequently, we investigated whether the cellular effects of IMP on CRPC were time-dependent. The results demonstrated that the inhibitory effects of IMP on PC3 and DU145 reached 48.76% and 34.06%, respectively, when IMP treatment was extended to 72 h (Fig. 1D).
Furthermore, we carried out a colony formation assay to determine the effect of IMP on the colony forming ability of CRPC cells. After 48 h of treatment, the number and size of colonies were significantly reduced with increasing IMP concentration in the IMP-treated group compared to the untreated group, suggesting that IMP inhibits the tumorigenic capacity of PC3 and DU145 cells in vitro (Fig. 1E and Figure S1B). This finding prompted us to assess the impact of IMP on the proliferative activity of PC3 and DU145 in greater depth. Due to the decreased cell proliferative activity, which results in the difficulty of incorporating EdU into DNA, we investigated the impact of IMP on the proliferative ability of PC3 and DU145 cells through the use of EdU (Radwan et al. 2022). The results demonstrated that the PC3 and DU145 cell lines exhibited a significantly reduced number of EdU-stained cells following treatment with high concentrations of IMP, in comparison to the control group (Fig. 1F, Figure S1C and S1D). Overall, these findings emphasize the effective inhibitory effect of IMP on the proliferation of PC3 and DU145 cells. In addition, we observed that the IMP was administered at a higher concentration, therefore we conducted a safety assessment of IMP. The results showed that the cell viability of RWPE-1 cells remained above 70% even at IMP concentrations up to 12 mM, indicating that IMP is less toxic to normal cells (Figure S1A). We also used Annexin-V-FITC/PI double staining and flow cytometry analysis to verify whether IMP induced apoptosis in PC3 and DU145 cells. Apoptosis in PC3 and DU145 cells did not change significantly after IMP treatment (Figure S1F). Taken together, these findings suggest that IMP has the ability to inhibit the proliferation of CRPC cells.
IMP inhibits the migration of PC3 and DU145To investigate the effects of IMP on CRPC migration, we use IMP treatment of PC3 and DU145 for cell migration experiments. In wound healing experiments, the healing area of cellular wounds decreased with increasing treatment concentrations. Specifically, at 10 mM, the healing rates of PC3 and DU145 cells were reduced to 44.19% and 6.13%, respectively (Fig. 1G, H and Figure S2A). Due to the inability of the scratch assay to effectively distinguish between cell migration and proliferation, we employed the Transwell migration assay for our studies. As shown in Fig. 1I and Figure S2B the number of cells migrating through the membrane decreased to varying extents following IMP treatment at concentrations of 1, 5, and 10 mM. This finding is consistent with the wound healing assay results, which demonstrated a reduction in the migratory ability of PC3 and DU145 cells after IMP treatment. Collectively, these results suggest that IMP inhibits migration of CRPC cells in a concentration-dependent manner.
Fig. 2
IMP could alter gene expression in PCa and result in the upregulation of PDZK1 expression. A Number of differentially expressed upregulated (log2FC ≥ 1 & q < 0.05) and downregulated (log2FC ≤ −1 & q < 0.05) genes in each comparison group. B Location of the PDZK1 gene in the volcano map. C The top 20 KEGG pathways with the smallest P-value (or Q-value) were utilized for mapping, with the vertical coordinate being the pathway name and the horizontal coordinate being the − log10 value of the P-value or Q-value of the KEGG pathway enrichment analysis for that KEGG pathway. D Histogram depicting mRNA expression levels in PC3 cells. E PDZK1 and NPR3 mRNA levels in PC3 cells. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001; ns, not significant
IMP exerts inhibitory effects on PCa cell phenotype via PDZK1We investigated the potential mechanisms by which IMP exerts its antiproliferative and antimigratory effects in CRPC cells. Transcriptome analysis revealed that IMP treatment significantly up-regulated 333 genes as well as significantly down-regulated 172 genes (Fig. 2A). In order to find the key genes, we performed an initial screen among the more significantly upregulated genes (|log2| (FC) > 1.5, trans_type is protein_codin and DO is associated with cancer). We screened eight genes including PDZK1 and explored them in the volcano map (Fig. 2B). Additionally, Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis revealed an enrichment of the PI3K-AKT signaling pathway (Fig. 2C). To identify genes with stable differences among the screened candidates, we further assessed the mRNA levels and ultimately selected the PDZK1 gene for detailed analysis (Fig. 2D, E). Increasing evidence suggests that PDZK1 plays a role in inhibiting tumor progression and enhancing sensitivity to chemotherapeutic agents, highlighting its potential as a promising therapeutic target (Ma et al. 2024; Qi et al. 2020; Wang et al. 2024). However, the impact of PDZK1 in PCa represents an intriguing emerging question. We first investigated PDZK1 levels in tumour tissue and normal tissue of PCa patients to study the potential role of PDZK1 in different tissues. tumour tissue exhibited significantly lower levels of PDZK1 protein compared with normal tissue (Figure S1E). Additionally, we evaluated the effect of IMP treatment on PDZK1 protein expression. Specifically, IMP treatment increased the expression level of PDZK1, suggesting that IMP may exert antiproliferative and antimigratory effects by promoting PDZK1 expression (Fig. 3A, B). However, no significant PDZK1 upregulation was observed in LNCap and 22RV1, which may explain the not effective of IMP on androgen-dependent prostate cancer cells (Fig. 3C, D).
Fig. 3
Knockdown of PDZK1 affects the proliferative ability of cells. A PDZK1 mRNA expression levels and protein expression levels in PC3. B PDZK1 mRNA expression levels and protein expression levels in DU145. C PDZK1 mRNA expression levels and protein expression levels in LNCap. D PDZK1 mRNA expression levels and protein expression levels in 22RV1. E Knockdown efficiency of PDZK1 in PC3. F Knockdown efficiency of PDZK1 in DU145. G Cell viability assay after PC3 knockdown of PDZK1. H Cell viability assay after DU145 knockdown of PDZK1. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001; ns: not significant
To assess the critical role of PDZK1 in PCa cell proliferation and migration, we employed small interfering RNA (siRNA) to knock down PDZK1 expression. Before investigating the role of PDZK1 in cells, we first assessed the efficiency of the siRNA-mediated knockdown. The results demonstrated that the knockdown of PDZK1 led to a significant reduction in both mRNA and protein levels in the cells (Fig. 3E, F). This suggests that siRNA is effective in knocking down PDZK1. Subsequently, we observed an increase in cell viability after knockdown of PDZK1 expression in the knockdown of PDZK1 cell viability assay (Fig. 3G, H). This suggests that PDZK1 may have the potential to inhibit cell proliferation.
Fig. 4
Knockdown of PDZK1 promotes PCa proliferation and migration. A Cell colony formation after PDZK1 knockdown in PC3 and DU145 cells. B Transwell migration assay after knockdown of PDZK1 in PC3 and DU145 cells. Scale bar, 200 μm. C Wound healing assay after knockdown of PDZK1 in PC3 cells. Scale bar, 500 μm. D Wound healing assay after knockdown of PDZK1 in DU145 cells. Scale bar, 500 μm. E Cell viability assay combining PDZK1 knockdown and IMP treatment using PC3. F Cell viability assay combining PDZK1 knockdown and IMP treatment using DU145. G Cell clones formed by PC3 and DU145 in a combined PDZK1 knockdown and IMP treatment experiment. H Transwell migration experiments of PC3 and DU145 in a combined PDZK1 knockdown and IMP treatment experiment. Scale bar, 200 μm. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001; ns, not significant
Additionally, cell cloning experiments demonstrated the impact of PDZK1 knockdown on colony formation, suggesting that PDZK1 plays a critical role in the proliferation of PC3 and DU145 cells (Fig. 4A and Figure S2C). To determine whether PDZK1 affects the migration of PC3 and DU145 cells, we conducted Transwell migration and wound healing assays following PDZK1 knockdown. PDZK1 knockdown resulted in increased cell migration (Fig. 4B-D and Figure S2D, E). These findings suggest that the expression level of PDZK1 plays a critical inhibitory role in the proliferation and migration of CRPC cells.
We defined the potential role of this gene in the process of IMP action through a combination of knockdown and drug treatment experiments. The results showed that si-PDZK + IMP treatment reduced the cell viability growth of si-PDZK-treated cells (Fig. 4E, F). It suggests that IMP may mediate the antiproliferative effect through PDZK1. In the colony formation assay of cells, we observed similar results to those of CCK8. Specifically, si-PDZK1 + IMP treatment increased the number of IMP-treated cell colonies (Fig. 4G and Figure S2F). In the Transwell migration, the si-PDZK1 + IMP group exhibited significantly reduced migration compared to the si-PDZK1 group (Fig. 4H and Figure S2G). In the wound healing rescue experiment, we observed similar results to those in the migration experiment (Fig. 5A, B). Taken together, these data suggest that IMP inhibits the proliferation and migration of PC3 and DU145 cells by upregulating PDZK1 expression.
Fig. 5
IMP influences PDZK1 protein and PI3K-AKT pathway protein levels in cells. A Wound healing assay results (left) and wound healing rate (right) of PC3 cells in a combined PDZK1 knockdown and IMP treatment experiment. Scale bar, 500 μm. B Wound healing assay results (left) and wound healing rate (right) of DU145 cells in a combined PDZK1 knockdown and IMP treatment experiment. Scale bar, 500 μm. C Representative western blot analysis of PC3 and DU145 cells with or without treatment with IMP (10 mM). D Representative western blot analysis of PC3 and DU145 cells with or without si-PDZK1 treatment. E PDZK1 protein expression levels in PC3 and DU145 cells treated with different concentrations of IMP. F Volumetric and gravimetric analysis of excised tumors (n = 5 per group). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001; ns: not significant
IMP inhibits activation of the PI3K-AKT signalling pathway by increasing PDZK1 expressionTranscriptomic analysis revealed that IMP treatment significantly regulated the phosphatidylinositol 3-kinase (PI3K)-protein kinase B (AKT) signalling pathway. Therefore, we examined the effect of IMP on the phosphorylation of this pathway. Specifically, IMP treatment reduced the phosphorylation level of the PI3K-AKT pathway in CRPC cells, suggesting that IMP exerts antiproliferative and antimigratory effects on CRPC by inhibiting the activation of the PI3K-AKT signalling pathway (Fig. 5C and Figure S3A). In addition, we evaluated the impact of PDZK1 knockdown on the phosphorylation of the PI3K-AKT pathway. In contrast to the results observed with IMP treatment, knockdown of PDZK1 led to hyperphosphorylation of the PI3K-AKT pathway (Fig. 5D and Figure S3B). In addition, we observed a progressive increase in PDZK1 protein levels following treatment of PC3 and DU145 cells with increasing concentrations of IMP (Fig. 5E and Figure S2H). This suggests that various concentrations of IMP contribute to the increased expression of PDZK1 and help maintain stable regulation of cell proliferation and migration throughout the experiment. It is well established that phosphorylation of the PI3K-AKT signaling pathway promotes the progression and poor prognosis of PCa. Taken together, these results demonstrate that IMP inhibits PI3K-AKT pathway activation by upregulating PDZK1 expression, which in turn suppresses the proliferation and migration of PC3 and DU145 cells.
Inhibition of subcutaneous graft tumor growth in nude mice by IMP Fig. 6
IMP inhibits tumor growth in vivo. A In vivo tumor photography and post-mortem tumor imaging after euthanasia of mice. B Pathological and immunohistochemical analysis of tumor tissues. Scale bar, 200 μm
To evaluate the in vivo anti-tumor efficacy and safety of IMP, we injected PC3 cells subcutaneously into the right axilla of mice. IMP was administered intraperitoneally every two days starting on the third day post-injection until the tumors became visibly apparent (Figure S3C). We observed that tumor volume and weight in the IMP-treated group were significantly reduced compared to the Control group (Figs. 5F and G and 6A). Additionally, we observed that IMP treatment preserved tumor tissue structure, reduced Ki-67 expression, and increased PDZK1 expression in the tumor tissue (Fig. 6B). Taken together, these findings suggest that IMP possesses significant antiproliferative and antimigratory potential and demonstrates safety in vivo.
留言 (0)