
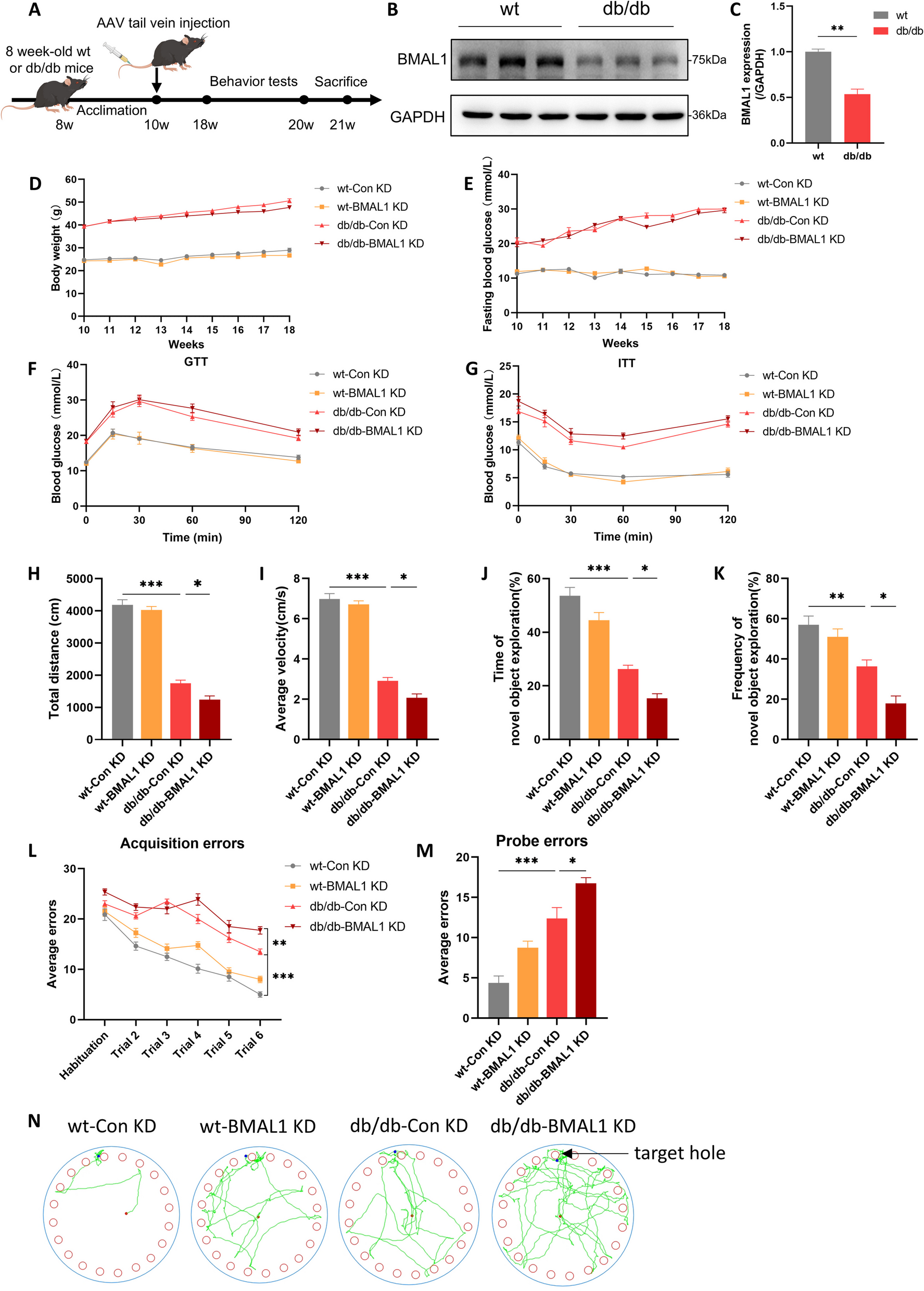
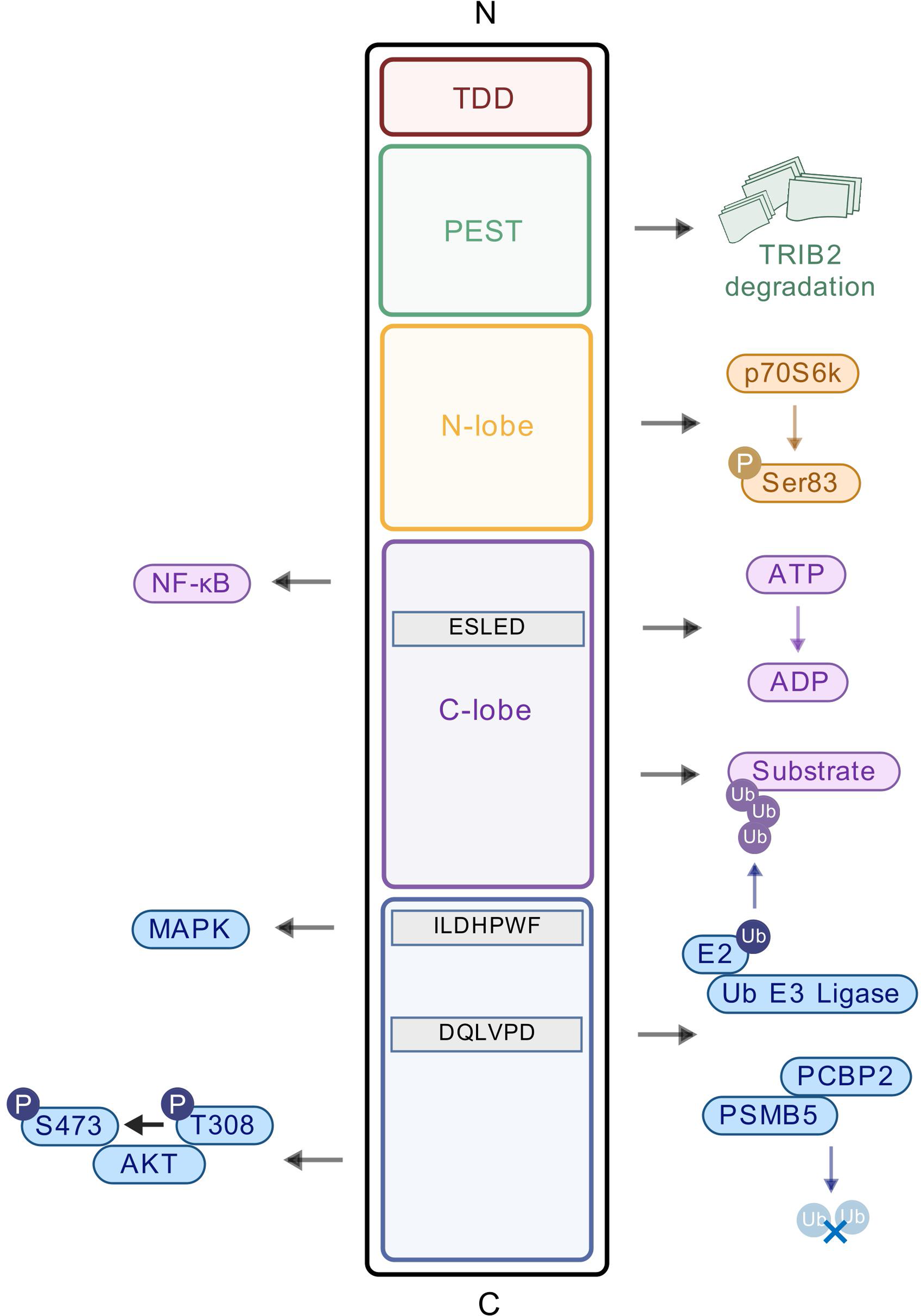
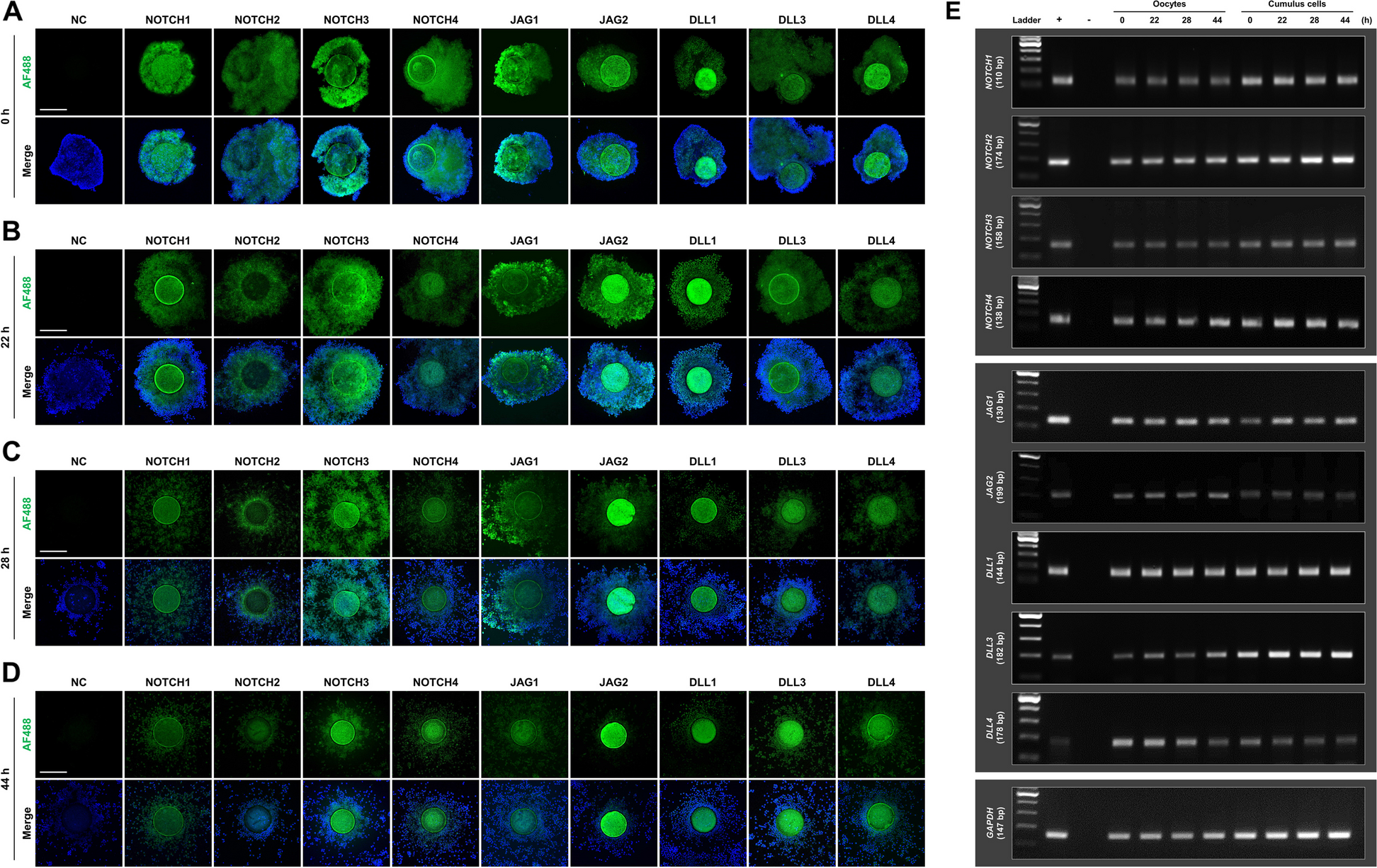
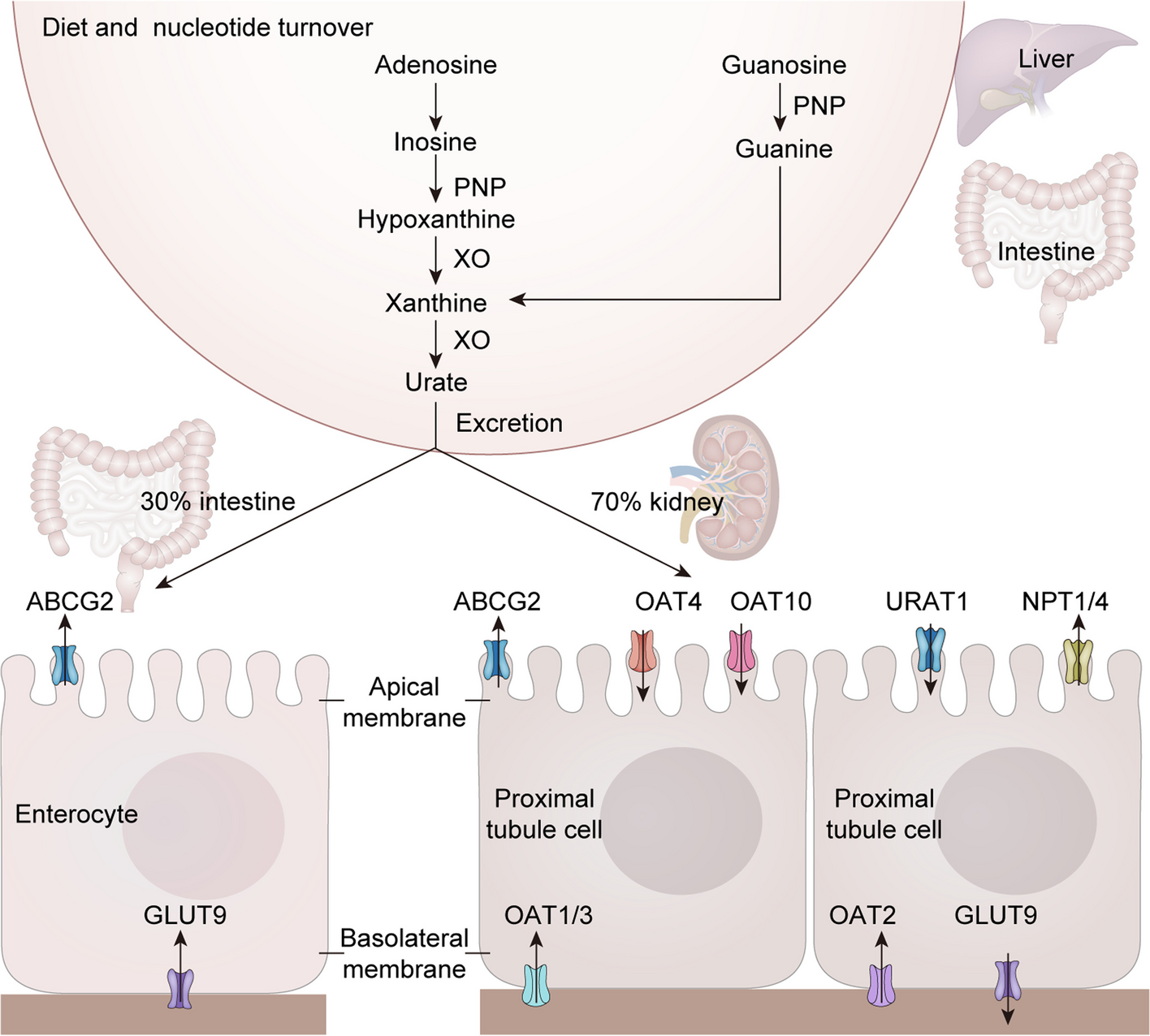
記住我




We first examined whether LRRK2 expressed in porcine oocytes, and as shown in Fig. 1A, the mRNA levels of LRRK2 during GV, GVBD, MI and MII stage were stable. There was no significant difference for the mRNA expression at different stages. Then we stained LRRK2 protein with an antibody and observed its localization in porcine oocytes. As shown in Fig. 1B, during meiotic maturation, LRRK2 mainly accumulated at the cortex area of porcine oocytes. Fluorescence intensity analysis indicated that the signals were also not change at different stages of meiotic maturation (Fig. 1C). These data suggested that LRRK2 existed in porcine oocytes and had specific localization pattern.
Fig. 1
Localization and expression of LRRK2 during porcine oocyte maturation. (A) The relative mRNA level of LRRK2 at GV, GVBD, MI and MII stage of porcine oocytes. There was no significantly difference between these groups. (B) Localization of LRRK2 at GV, GVBD, MI and MII stage of porcine oocytes. LRRK2 mainly accumulated at the cortex area of porcine oocytes. Red, LRRK2; blue, DNA. Bar = 30 μm. (C) The fluorescence intensity analysis of LRRK2 protein by antibody staining. There was no significantly difference between these groups
Loss of LRRK2 activity induces meiotic maturation defects in porcine oocytesSince LRRK2 expressed in porcine oocytes, we then explored LRRK2-IN-1 treatment to inhibit the LRRK2 activity to explore the potential roles of LRRK2 during meiotic maturation. As shown in Fig. 2A, the control cumulus-oocyte complexes (COCs) showed expanded cumulus cells, a marker for the oocyte maturation. While we found that different dose with 10 µM, 20 µM and 40 µM of LRRK2-IN-1 all caused the cumulus expansion defects of COCs. To further confirmed the effects of LRRK2 on porcine oocytes, we also examined the polar body extrusion, the direct evidence for oocyte maturation, and the statistical analysis data indicated that loss of LRRK2 activity disturbed the rate of polar body extrusion rate, and this was dose-dependent effects (Control, 68.98% ± 2.04%, n = 100; 10 µM group: 46.59% ± 1.10%, n = 100, P < 0.001; 20 µM group: 37.39% ± 4.65%, n = 100, P < 0.01; 40 µM group: 6.17% ± 0.67%, n = 100, P < 0.001) (Fig. 2B). These data suggested that LRRK2 activity was essential for porcine oocyte maturation. 20 µM of LRRK2-IN-1 was adopted for the following investigations.
Fig. 2
Loss of LRRK2 activity on cumulus cell expansion and polar body extrusion of porcine oocytes. (A) 10 µM, 20µM and 40µM dose of LRRK2-IN-1 treatment all caused the failure of cumulus cell expansion. Bar = 200 μm. (B) 10 µM, 20µM and 40µM dose of LRRK2-IN-1 treatment all caused the failure of polar body extrusion of porcine oocytes. The statistical analysis data showed the significantly difference between treatment group and control group. **, P < 0.01; ***, P < 0.001
Loss of LRRK2 activity disrupts cytoskeleton dynamics in porcine oocytesTo explore the potential mechanism of LRRK2 on porcine oocyte meiotic maturation, we first examined the spindle organization and chromosome alignment. As shown in Fig. 3A, the control oocytes at MI stage showed normal barrel-shape meiotic spindle, with well alignment chromosomes, however, inhibition of LRRK2 activity caused the failed formation of spindle, the microtubules accumulated around the chromosomes, while the chromosomes also showed aberrant accumulation or mis-alignment. The rate of aberrant spindle formation by statistical analysis data in the treatment group was significantly higher than the control group (Control, 27.8% ± 6.4% n = 69 vs. treatment 52.3% ± 4.2%, n = 71, P < 0.01) (Fig. 3B); similarly, the rate of misaligned chromosomes was also significantly higher than the control group (Control, 30.5% ± 4.8% n = 69 vs. treatment 62.7% ± 9.5%, n = 71, P < 0.01) (Fig. 3C). We then examined the actin filaments distribution, and as shown in Fig. 3D, we found that the fluorescence signals were both decreased at both cytoplasmic actin and cortex actin. The fluorescence intensity analysis showed that actin in the cytoplasm significantly decreased compared with the control group (1 vs. 0.20 ± 0.07, n = 30, P < 0.001) (Fig. 3E), and similar results were also found for the cortex actin (1 vs. 0.61 ± 0.15, n = 30, P < 0.01) (Fig. 3E). Actin is the main power for the spindle movement, and we then examined the spindle position, and the results showed that the spindles were not attached to the cortex in most oocytes after LRRK2 inhibition (Fig. 3F). The measurement of chromosome-cortex distance also confirmed this finding (3.97 μm ± 1.93 μm vs. 17.90 μm ± 4.62 μm, n = 30, P < 0.05) (Fig. 3G). These data suggested that LRRK2 activity was essential for cytoskeleton dynamics during porcine oocyte maturation.
Fig. 3
Loss of LRRK2 activity disturbs spindle and actin dynamics of porcine oocytes. (A) LRRK2-IN-1 treatment caused the spindle formation defect and chromosome misalignment of porcine oocytes. Control oocytes showed barrel shape spindle and well-aligned chromosomes; treated oocytes showed aberrant microtubule and chromosome accumulation. Green, microtubule; blue, DNA. Bar = 30 μm. (B) The statistical analysis data showed the significantly difference of the percentage of aberrant spindle between treatment group and control group. **, P < 0.01. (C) The statistical analysis data showed the significantly difference of the percentage of misalignment chromosomes between treatment group and control group. **, P < 0.01. (D) LRRK2-IN-1 treatment caused the aberrant distribution of cytoplasmic actin and cortex actin in porcine oocytes. Actin signals decreased in both cytoplasm and cortex of oocytes. Red, actin; blue, DNA. Bar = 30 μm. (E) Relative fluorescence intensity analysis of cytoplasmic actin and cortex actin showed a significant decrease after LRRK2-IN-1 treatment in porcine oocytes. **, P < 0.01; ***, P < 0.001. (F) The distance of chromosomes to the oocyte cortex was much longer in the LRRK2 treatment group compared with the control group. Red, DNA. Bar = 30 μm. (G) The measurement of chromosome-cortex distance in the control and LRRK2-inhibited oocytes. *, P < 0.05
LRRK2 associates with mitochondria for its distribution in porcine oocytesIn porcine oocytes, mitochondria accumulate at the cortex during meiotic maturation, while LRRK2 localization showed similar pattern in our results. To confirm this hypothesis, we co-stained LRRK2 and mitochondria, and the results showed that LRRK2 co-localized with mitochondria at metaphase I stage of porcine oocytes (Fig. 4A), indicating the potential link of each other. The subsequent data showed that loss of LRRK2 activity caused the aberrant accumulation of mitochondria of porcine oocytes: control oocytes showed continuous and uniformly distribution of mitochondria at the cortex area, however, the mitochondria showed clustered aberrant accumulation and sporadic distribution at the cortex of oocytes with loss of LRRK2 activity (Fig. 4B). The rate of aberrant mitochondria distribution was much higher in the treatment group compared with the control group (Control, 23.6% ± 4.9%, n = 54 vs. treatment 75.3% ± 7.3%, n = 46, P < 0.01); while the intensity of mitochondria signals in the treated oocytes was also much higher than the control oocytes (1 vs. 3.2 ± 0.48, n = 30, P < 0.001). These data suggested that LRRK2 was linked with mitochondria and its activity determines the distribution of mitochondria in porcine oocytes.
Fig. 4
Loss of LRRK2 activity affects mitochondria distribution of porcine oocytes. (A) Co-staining of LRRK2 and mitochondria showed that these two co-localize with each other at the metaphase I of porcine oocytes. We first stained Mito-Tracker in alive oocytes, and then fixed the oocytes for the LRRK2 antibody staining. Green, LRRK2; red, mitochondria; blue, DNA. Bar = 30 μm. (B) LRRK2-IN-1 treatment caused the mitochondria distribution defect of porcine oocytes. Mitochondria accumulated at the cortex area of control oocytes, while mitochondria showed clustered distribution after inhibition of LRRK2 activity. Red, mitochondria. Bar = 30 μm. (C) The statistical analysis data showed the significantly increase for the percentage of aberrant mitochondria distribution between treatment group and control group. ***, P < 0.001. (D) Relative fluorescence intensity analysis of mitochondria showed a significant increase after LRRK2-IN-1 treatment in porcine oocytes. **, P < 0.01
Loss of LRRK2 activity induces oxidative stress in porcine oocytesTo further analyze the association of LRRK2 with mitochondria, we then examined the mitochondria membrane potential (MMP) and the following ROS level. Since MMP reflects the functions of mitochondria and ROS level is determined by mitochondria function. Our results showed that the TMRE level increased after inhibition of LRRK2 activity: there was barely signals of TMRE in the control oocytes, while strong signals concentrated at the cortex were observed in the treated oocytes (Fig. 5A), and the fluorescence intensity analysis data also confirmed this (1 vs. 3.37 ± 0.33, n = 30, P < 0.01), indicating the mitochondria dysfunction after loss of LRRK2 activity. Meanwhile, the ROS level was also increased in the treated oocytes: most oocytes showed positive ROS signals while there were barely oocytes showing with ROS signals in the control group (Fig. 5C), and the fluorescence intensity of ROS in the treated oocytes was significantly higher than the control oocytes (1 vs. 2.45 ± 0.37, n = 30, P < 0.01). These data suggested that LRRK2 inhibition induced mitochondria dysfunction and further caused oxidative stress in porcine oocytes.
Fig. 5
Loss of LRRK2 activity induces oxidative stress in porcine oocytes. (A) LRRK2-IN-1 treatment caused the increase of TMRE fluorescence signals in porcine oocytes. Red, TMRE. Bar = 30 μm. (B) The statistical analysis data showed the significantly increase of TMRE fluorescence intensity in the treatment group compared with the control group. **, P < 0.01. (C) LRRK2-IN-1 treatment caused the increase of ROS fluorescence signals in porcine oocytes. Green, ROS. Bar = 150 μm. (D) The statistical analysis data showed the significantly increase for the ROS signals after LRRK2-IN-1 treatment. **, P < 0.01
留言 (0)