
記住我

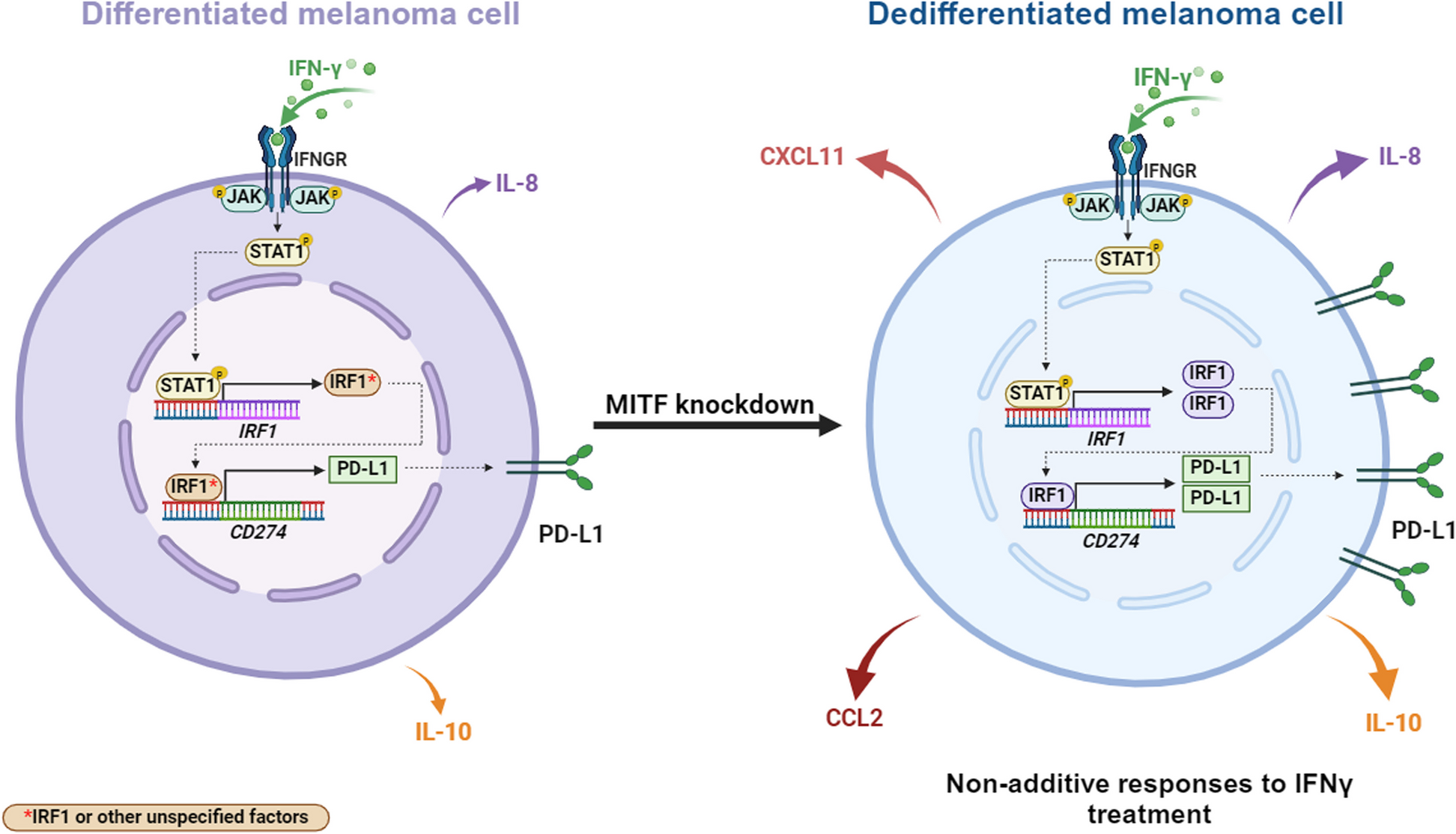








Dedifferentiated melanoma cells have been reported to display enhanced responses to IFNγ stimulation when compared to differentiated melanoma cells. This response is characterized by an increase in both the expression of interferon response genes and access to chromatin regions [4]. Since IFNγ induces both the expression of immunostimulatory and immunosuppressive genes [27], we set out to investigate the potential effect of melanoma dedifferentiation on IFNγ-induced gene expression, particularly on immune related genes. To do that, we induced dedifferentiation in 624Mel melanoma cells via siRNA mediated MITF knockdown and treated the cells with IFNγ and then performed bulk RNA sequencing on both differentiated (siCTRL) and dedifferentiated (siMITF) 624Mel cells, with or without IFNγ treatment. Treatment with siMITF resulted in significant downregulation of MITF itself and multiple MITF target genes, e.g., MLANA, DCT, PMEL and TRPM1 (Fig. 1A). Similarly, treatment with IFNγ induced expression of a multitude of known IFN response genes, e.g., STAT1, STAT3, IRF1, CXCL11, IDO1, HLA-E and HLA-C (Fig. 1A). This collectively indicates that the siMITF and IFNγ treatments independently produce their intended effects in the 624Mel cells. Principal component analysis (PCA) on substantially expressed genes (top 2,000 expressed genes) confirmed that the siCTRL and siMITF treated 624Mel cells have distinct transcriptional states at baseline, and when treated with IFNγ (Fig. 1B). Notably, a subset of 320 genes respond only to the concomitant treatment of MITF knockdown and IFNγ treatment (Fig. 1C). To further characterize the effect of MITF knockdown and IFNγ treatment on the differentiation status of 624Mel cells, we performed GSEA using established gene sets that specifically associate with differentiation stages in melanoma cells (Tsoi et al. 2018)[8]. Irrespective of IFNγ treatment, the siMITF treated cells showed increased expression of the “Undifferentiated” and “Neural-Crest Like” gene sets, whereas the “Melanocytic” and “Transitory” sets were reduced compared to the siCTRL treated cells, indicating that knocking down MITF expression is sufficient to induce dedifferentiation (Fig. 1C, Supplementary Fig. S1).
Fig. 1
MITF knockdown induces dedifferentiation in 624Mel melanoma cells. A Volcano plots of differentially expressed genes, showing the independent effects of siMITF (top) and IFNγ (bottom) on 624Mel cells. B PCA of transcriptome profiles according to different experimental conditions. C Overlap of differentially expressed genes depending on responses to either MITF knockdown or IFNγ treatment, or to the concomitant factors of MITF knockdown and IFNγ treatment. D GSEA comparing the expression of the Tsoi differentiation gene sets in siMITF treated cells compared to the siCTRL treated cells
To further elucidate how dedifferentiation affects the IFNγ response in melanoma cells, we applied a directional interaction classification system, conceptualized by Piggott et al. (2015) [41], and gene ontology enrichment analysis to our quantified transcriptional profiles. We identified a total of 148 genes showing significant non-additive responses upon combined treatment of siMITF and IFNγ (likelihood ratio test, corrected P < 0.05). This response emerges from genes that respond to neither factor (Fig. 2A), from genes that respond to IFNγ only (Fig. 2B), siMITF only (Fig. 2C) or both factors, but whose concomitant response is different from the sum of responses to each factor (Fig. 2D). Importantly, in all cases except for the siMITF single effect responders, the directional interaction of IFNγ and MITF knockdown was predominantly characterized by positive synergism, with non-additive responders showing higher expression than the sum of single effects of IFNγ and MITF knockdown. After functional enrichment analysis on this non-additive response gene set, eight out of 13 enriched GO terms represent cellular processes implicated in adaptive immunity: “T cell proliferation”, “MHC protein complex II”, “Immune response”, “MHC protein complex”, “Antigen processing and presentation”, “Cytokine mediated signaling”, “Regulation of T cell activation” and “Response to cytokine”. Furthermore, an analysis of transcription factor target enrichment revealed increased transcriptional activity for several of the JAK/STAT pathway components, namely STAT1, STAT3, IRF1, IRF2 and CIITA. Notably, among the non-additive response genes was CD274 (Fig. 2B), the gene encoding for PD-L1. These results collectively indicate that dedifferentiation drives melanoma cells into a distinct cellular state. Upon IFNγ stimulation, this state leads to a synergistic increase in the expression of immune-related genes through enhanced JAK/STAT signaling.
Fig. 2
Dedifferentiation and IFNγ stimulation synergistically affect expression of immune related genes in 624Mel melanoma cells. A Non additive response emerging from single-effect non-responders (52 genes total). B Non additive response emerging from IFNγ responders (36 genes total). C Non additive response emerging from siMITF responders (49 genes total). D Non additive response emerging from IFNγ & siMITF responders (11 genes total)
Dedifferentiated 624Mel melanoma cells simulated with IFNγ secrete greater quantities of multiple immunomodulatory cytokinesDedifferentiated melanoma cells have been reported to secrete multiple cytokines in greater quantity, notably CCL2, CCL5 and IL-1β, either without inflammatory stimuli or when stimulated with TNFα [2, 18]. As our RNAseq data showed a synergistic transcriptional response on cytokine expression by IFNγ stimulated dedifferentiated 624Mel cells, we investigated the secretome of dedifferentiated 624Mel cells by measuring a panel of 18 cytokines and chemokines using the Luminex Discovery assay. Out of the 18 analytes, 13 were detected. CCL2 and IL-10 secretion was significantly increased (one-way ANOVA, P < 0.0001) in dedifferentiated 624Mel cells upon IFNγ treatment (Fig. 3A). Notably, a single value for CCL2 in the siMITF + IFNγ group measured above the assays standard curve and was thus not included in the statistical analyses. CXCL10 and PD-L1 also displayed a synergistic pattern (Fig. 3B). However, the expression values for CXCL10 and PD-L1 were extrapolated beyond the standard curves of the assay, with CXCL10 values for the siMITF + IFNγ group measuring above the standard curve whilst all values for PD-L1 measured below it. Additionally, secretion of IL-8, CXCL11 and CXCL1 was increased in dedifferentiated 624Mel cells, with IL-8 and CXCL11 being significantly increased but not CXCL1 (one-way ANOVA, P < 0.0001, P = 0.084 for CXCL1) (Fig. 3C). However, these analytes did not show a pattern of synergistic increase upon IFNγ treatment (Fig. 3C). Lastly, VEGF secretion was detected in all groups but did not differ significantly in any of the experimental conditions (Fig. 3D). Other detected analytes were mostly extrapolated below the standard curves of the assay (data not shown due to values falling below the lower limits of detection). These results indicate that dedifferentiation predisposes 624Mel melanoma cells to increase IFNγ-induced cytokine secretion.
Fig. 3
Dedifferentiated 624Mel melanoma cells stimulated with IFNγ secrete greater quantities of multiple immunomodulatory cytokines. A Luminex assay measuring secretion (pg/mL/106 cells) of CCL2, IL-10 in 624Mel cells transfected with either siCTRL or siMITF, with or without IFNγ. B Luminex measurements for CXCL10 and PD-L1 secretion in 624Mel cells, same conditions as in (A). C Luminex measurements for IL-8, CXCL11 and CXCL1 secretion in 624Mel cells, same condition as in (A). D Luminex measurement for VEGF secretion in 624Mel cells, same condition as in (A). Plots indicate means along with ranges from minimum to maximum values. Statistical analysis performed by one-way ANOVA and Tukey’s multiple comparisons test, adjusted P value * = < 0.05, ** = < 0.01, *** = < 0.001, **** = < 0.0001 (n = 3, for CCL2 in the siMITF + IFNγ group, n = 2)
Synergistic increase in PD-L1 expression in dedifferentiated 624Mel melanoma cells upon IFNγ stimulationSince PD-L1 is a clinically relevant immunotherapy marker [45], we decided to further investigate this pattern of synergistic increase in PD-L1 expression. As shown in Fig. 4A-D, siRNA mediated MITF knockdown in 624Mel melanoma cells in combination with IFNγ (5 ng/mL) treatment significantly increased PD-L1 expression at both the RNA (Fig. 4B; one-way ANOVA, P < 0.01) and protein levels (Fig. 4D; one-way ANOVA, P < 0.001) compared to the expected increase caused by IFNγ treatment alone. Critically, without IFNγ, MITF knockdown led to a non-significant decrease in PD-L1 mRNA expression whereas protein expression remained unchanged.
Fig. 4
Dedifferentiated 624Mel melanoma cells express more PD-L1 upon IFNγ stimulation. Quantitative RT-PCR of MITF (A) and PD-L1 (B) mRNA expression in 624Mel cells transfected with Control siRNA (siCTRL) or siMITF, with or without 5 ng/mL IFNγ. C, E Western blotting of MITF and β-Actin as control in 624Mel cells transfected with siCTRL or siMITF, with or without 5 ng/mL IFNγ. D, F FACS analysis of PD-L1 protein expression on 624Mel cells transfected with siCTRL or siMITF, with or without 5 ng/mL IFNγ. Plots indicate mean ± standard deviations of value distributions. Statistical analysis performed by one-way ANOVA and Tukey’s multiple comparisons test, adjusted P value * = < 0.05, ** = < 0.01, *** = < 0.001, **** = < 0.0001 (n = 5 for A & B, n = 3 for C & D)
Next, we investigated the generalizability of this pattern of PD-L1 expression. Contrary to our expectations, when we analyzed the effects of MITF/Mitf knockdown on the IFNγ/Ifnγ-induced PD-L1/Pd-l1 mRNA expression in additional melanoma cell lines, including two mouse melanoma cell lines (B16 and YUMM1.7) for interspecies comparison, we did not observe the same effects as seen in the 624Mel melanoma cell line. In fact, MITF/Mitf knockdown followed by IFNγ/Ifnγ treatment resulted in reduced PD-L1/Pd-l1 mRNA expression in the human A375P and mouse B16 cells lines, whereas Ifnγ treatment significantly increased Mitf expression in the mouse YUMM1.7 cells (Fig. 5). Similarly, MITF knockdown in Malme-3M cells did not result in increased PD-L1 protein expression following IFNγ treatment (Supplementary Fig. S3). This clearly indicates that increased PD-L1 expression following IFNγ treatment and MITF knockdown, as well as the crosstalk between IFNγ and MITF expression in melanoma cells, is context-dependent.
Fig. 5
IFNγ/Ifnγ-induced PD-L1 mRNA expression is not synergistic following MITF/Mitf knockdown in SK-MEL-28, A375P, B16 and YUMM1.7 melanoma cells. Graphs showing the relative expression of MITF/Mitf and PD-L1/Pd-l1 as determined by qPCR in the indicated cell lines upon MITF/Mitf knockdown and IFNγ treatment. Plots indicate mean ± standard deviations of value distributions. Statistical analysis performed by one-way ANOVA and Tukey’s multiple comparisons test, adjusted P value * = < 0.05, ** = < 0.01, *** = < 0.001, **** = < 0.0001 (n = 3)
IFNγ-induced PD-L1 expression in 624Mel melanoma cells is dependent on JAK activity irrespective of differentiation stateSince IFNγ-induced PD-L1 expression is mediated through the JAK1/2-STAT1/3-IRF1 axis in melanoma cells [36], and since IRF1 appeared among the most significantly affected genes in our RNA sequencing analysis (Fig. 2B), we investigated whether this non-additive increase in PD-L1 expression was dependent on the JAK-STAT1-IRF1 pathway. We began by applying a JAK inhibitor to our experiments in addition to the IFNγ treatment. The JAK kinases are immediately downstream of the IFNγ receptor complex and upon activation the JAKs phosphorylate the STAT transcription factors, inducing the expression of interferon stimulated genes (ISGs) [46,47,48]. As seen in Fig. 6A-J, inhibiting JAK activity significantly decreased STAT1 phosphorylation (Fig. 6D & F; one-way ANOVA, P < 0.0001), leading to decreased IRF1 mRNA and protein expression (Fig. 6B, G & I) and complete abrogation of IFNγ-induced PD-L1 expression in 624Mel cells, irrespective of MITF expression (Fig. 6C, G & J).
Fig. 6
IFNγ-induced PD-L1 expression in 624Mel cells is dependent on JAK kinase activity irrespective of differentiation. A, B, C Quantitative RT-PCR of MITF, IRF1 and PD-L1 mRNA expression in 624Mel cells transfected with control siRNA (siCTRL) or siMITF, with or without 5 ng/mL IFNγ and/or 300 nM JAK inhibitor. D, E, F Western blotting of STAT1, pSTAT1 (Tyr701) and β-Actin in 624Mel cells transfected with siCTRL or siMITF, with or without 5 ng/mL IFNγ and/or 900 nM JAK inhibitor. G, H, I, J Western blotting of MITF, IRF1, PD-L1 and β-Actin in 624Mel cells transfected with siCTRL or siMITF, with or without 5 ng/mL IFNγ and/or 900 nM JAK inhibitor. Plots indicate mean ± standard deviations of value distributions. Statistical analysis performed by one-way ANOVA and Tukey’s multiple comparison test, adjusted Pvalue ** = < 0.01, *** = < 0.001, **** = < 0.0001 (n = 5 for A-C, n = 4 for E-J)
IFNγ-induced PD-L1 expression in 624Mel melanoma cells is dependent on STAT1 irrespective of differentiation stateSince high IFNγ-induced PD-L1 expression in dedifferentiated 624Mel cells is dependent on JAK activity, we next wanted to identify which of the STAT members downstream of JAK is involved specifically. It has been reported that STAT1 and STAT3 are the main regulators of IFNγ-induced PD-L1 expression [36]. As seen in Fig. 7A-I, siRNA-mediated knockdown of STAT1 led to significantly decreased IRF1 expression (Fig. 7C & H, one-way ANOVA, P < 0.001) and complete abrogation of IFNγ-induced PD-L1 expression in 624Mel cells, irrespective of MITF expression (Fig. 7D & I). This is similar to the results seen when applying the JAK inhibitor (Fig. 6J). Whilst NF-κB inhibition led to reduced MITF expression in 624 Mel cells, inhibition of neither STAT3 nor NF-κB impacted PD-L1 expression in IFNγ-induced cells. (Supplementary Fig. S4).
Fig. 7
IFNγ-induced PD-L1 expression is dependent on STAT1, irrespective of differentiation status. A, B, C, D Quantitative RT-PCR of MITF, STAT1, IRF1 and PD-L1 mRNA expression in 624Mel cells transfected with siCTRL, siMITF, siSTAT1 or both siMITF and siSTAT1, with or without 5 ng/mL IFNγ. E, F, G, H, I Western blotting of MITF, STAT1, IRF1, PD-L1 and β-Actin in 624Mel cells transfected with siCTRL, siMITF, siSTAT1 or both siMITF and siSTAT1 with or without 5 ng/mL IFNγ. Plots indicate mean ± standard deviations of value distributions. Statistical analysis performed by one-way ANOVA and Tukey’s multiple comparisons test, adjusted P value ** = < 0.01, *** = < 0.001, **** = < 0.0001 (n = 3)
High IFNγ-induced PD-L1 expression in dedifferentiated 624Mel melanoma cells is dependent on IRF1Knowing that 624Mel cells are dependent on STAT1 for IFNγ-induced PD-L1 expression, we next examined the role of IRF1 in mediating the high PD-L1 expression in dedifferentiated 624Mel cells. First, we observed that siRNA mediated double knockdown of MITF and IRF1 resulted in reduced mRNA levels of both factors (Fig. 8A-B & Fig. 8D-E). Furthermore, the expression of IRF1 and PD-L1 was synergistically increased in the dedifferentiated 624Mel cells upon IFNγ treatment (Fig. 8E-F). Surprisingly, IFNγ-induced PD-L1 mRNA expression was significantly increased in the siIRF1 group compared to the IFNγ treated control (Fig. 8C; one-way ANOVA, P < 0.0001). Notably, this increase was not observed at the protein level, as IFNγ-induced PD-L1 protein expression was not significantly affected by siIRF1 knockdown alone (Fig. 8F). However, double knockdown of MITF and IRF1 decreased IFNγ-induced PD-L1 mRNA and protein expression in the dedifferentiated 624Mel cells, resulting in similar expression levels to that of the IFNγ treated control cells (Fig. 8C & F). This indicates that IRF1 is involved in mediating the high PD-L1 expression observed in IFNγ stimulated dedifferentiated 624Mel cells.
Fig. 8
IFNγ-induced PD-L1 expression in dedifferentiated melanoma cells is dependent on IRF1. A, B, C Quantitative RT-PCR of MITF, IRF1 and PD-L1 mRNA expression in 624Mel cells transfected with Control siRNA (siCTRL), siMITF, siIRF1 or both siMITF and siIRF1, with or without 5 ng/mL IFNγ. E, F, G Western blotting of MITF, STAT1, IRF1, PD-L1 and β-Actin in 624Mel cells transfected with siCTRL, siMITF, siIRF1 or both siMITF and siIRF1, with or without 5 ng/mL IFNγ. Plots indicate mean ± standard deviations of value distributions. Statistical analysis performed by one-way ANOVA and Tukey’s multiple comparisons test, adjusted P value * = < 0.05, ** = < 0.01, *** = < 0.001, **** = < 0.0001 (n = 4 for A-C, n = 5 for D-G)
Patient derived melanoma cell lines recapitulate a context-dependent relationship between dedifferentiation and IFNγ responsesAs our results indicate that dedifferentiation affects IFNγ-induced PD-L1 expression in a context-dependent manner in 624Mel cells and is not observed in our other melanoma cell lines (Fig. 4, Supplementary Fig. S3), we asked the question whether this trend is noticeable in a wider context of melanoma cases. Publicly available RNA sequencing data (GSE154996) consisting of 45 patient derived melanoma cell lines, either with or without 6-h treatment with 5 ng/mL IFNγ. Whilst PD-L1 mRNA expression is highly associated with the overall IFNγ response in the dataset (Fig. 9A, Spearman rs = 0.703, P < 0.0001) there is also a weak negative association between MITF and IFNγ-induced PD-L1 mRNA expression (Fig. 9A, Spearman rs = -0.3956, P = 0.0072) (Supplementary Fig. S7) [33]. Similarly, ranking these melanoma cell lines by MITF expression (Fig. 9B) revealed significantly higher PD-L1 expression among cell lines with lower MITF levels (Fig. 9C; Mann–Whitney U test, P < 0.05), indicating that whilst not universal, there is a tendency for higher IFNγ-induced PD-L1 expression among less differentiated melanoma cells.
Fig. 9
Dedifferentiated melanoma cell lines have higher IFNγ-induced PD-L1 expression and increased immune related gene expression. A Scatter plot and Spearman correlation analysis for association of PD-L1 mRNA expression with IFNγ response and MITF expression in 45 patient derived melanoma cell lines. B, C MITF and PD-L1 expression of IFNγ treated melanoma cell lines when divided by MITF expression. D The top quartile (MITFhigh) and bottom quartile (MITFlow) melanoma cell lines ranked by MITF expression. E Principal component analysis of MITFhigh and MITFlow melanoma cells, with or without IFNγ treatment. F PD-L1 expression of the MITFhigh and MITFlow melanoma cell lines. G GSEA of immunologically associated hallmark gene sets upregulated in IFNγ treated MITFlow melanoma cells compared to IFNγ treated MITFhigh melanoma cells. Lines in scatterplots represent medians. Statistical analysis performed by Spearman correlation analysis (A) and Mann–Whitney U test (B, D & F), P value * = < 0.05, **** = < 0.0001 (n = 22 for upper 50%, n = 23 for Lower 50%)
To further investigate what may facilitate this difference in PD-L1 expression, we compared the top and bottom quartiles of the melanoma cell lines, ranked by MITF expression (Fig. 9D). These two groups had clearly distinct transcriptional states (Fig. 9E), with the MITFhigh cells appearing differentiated and the MITFlow cells displaying a more dedifferentiated phenotype according to their expression of the Tsoi differentiation signatures (Supplementary Fig. S8).Both groups had appreciable variation in PD-L1 expression. However, most of the cells with high IFNγ-induced PD-L1 expression belonged to the MITFlow group (Fig. 9F; Supplementary Fig. S14A), although the difference in median PD-L1 expression did not prove statistically significant (Mann–Whitney U test, P = 0.16). Additionally, the IFNγ treated MITFlow cells displayed significantly higher expression of CCL2 and IL-8 (Kruskal Wallis test, P < 0.01) (Supplementary Fig. 14B & D). Additionally, CCLE2 mRNA expression was significantly increased in the MITFlow group upon IFNγ treatment (two-tailed paired Wilcoxon test, P = 0.0024) (Supplementary Fig. S14C), whilst IL-8 mRNA expression was not significantly affected by IFNγ (Supplementary figure). Similarly, the median expression of CXCL10 and CXCL11 was higher in the MITFlow cells, but the difference was not statistically significant (Mann–Whitney U test, P = 0.0780 for CXCL10 and P = 0.1561 for CXCL11) (Supplementary Fig. S14F). However, IL-10 was not widely expressed in neither group of melanoma cell lines (Supplementary Fig. S14G). Gene set enrichment analysis of the hallmark signature gene sets revealed that several immunological gene sets were significantly upregulated in the IFNγ treated MITFlow group compared to the IFNγ treated MITFhigh group (Fig. 9G). However, the IFNγ response signature gene set was not among the most highly significant gene sets, although it appeared upregulated in the MITFlow group (Supplementary Fig. S9). Overall, these results indicate that dedifferentiated melanoma cells have a widely enhanced response to IFNγ compared to their differentiated counterparts, resulting in greater expression of a multitude of immunologically relevant genes that are not exclusive to the IFNγ response. This supports our findings in the 624Mel cell line (Fig. 2) and further associates high IFNγ-induced PD-L1 expression with dedifferentiated melanoma cells.
To explain the molecular context of high PD-L1 expression in the dedifferentiated melanoma cells, we divided the MITFlow group into PD-L1high and PD-L1low based on their IFNγ-induced PD-L1 expression (Supplementary Fig. S10A) and then compared those two groups. Based on their overall mRNA expression, correlation between the individual MITFlow cell lines ranged from -0.38 to 0.16 and was non-significant in all cases (Supplementary Fig. S10B). However, minor inherent differences in the transcriptional states of the PD-L1high and PD-L1low groups were detected (Fig. 10A). Furthermore, gene set enrichment analysis revealed that the PD-L1high cells have significantly upregulated immunological gene signatures (Fig. 10B), with the top five most highly upregulated hallmark gene sets all belonging to inflammatory signaling, one of which being the IFNγ response signature gene set (Fig. 10C). To identify key genes impacting the PD-L1high group’s gene set signature profile, we conducted a leading-edge subset analysis [42] on the top five most significant gene sets. We identified a core set of 20 genes present in at least three leading-edge subsets (Fig. 10D). Notably, IRF1 was common to all subsets, indicating its strong influence on the gene set signature profile of the PD-L1high group. Altogether, these results indicate that dedifferentiated melanoma cells tend to be more responsive to IFNγ stimulation, and that high IFNγ-induced PD-L1 expression among dedifferentiated melanoma cells is associated with a broad enhancement of inflammatory signaling where IRF1 expression is a central contributing factor.
Fig. 10
Dedifferentiated melanoma cells with high IFNγ-induced PD-L1 expression are characterized by increased inflammatory signalling that converges on IRF1 expression. A PCA of MITFlow melanoma cell by high or low IFNγ-induced PD-L1 expression, along PC3 and PC5. B Top significantly upregulated hallmark gene sets in the MITFlow/PD-L1high group compared to the MITFlow/PD-L1low group. E GSEA plot showing enrichment of the IFNγ response gene set in MITFlow/PD-L1high melanoma cells. D Leading edge analysis defining the core set of genes contributing to the enrichment of the top five most significantly upregulated hallmark gene sets in the MITFlow/PD-L1high group
A similar comparison within the MITFhigh group (Supplementary Fig. S12A) revealed that high PD-L1 expression among the MITFhigh cells was also associated with enrichment of multiple inflammatory gene sets compared to MITFhigh cells with low PD-L1 expression (Supplementary Fig. S12B). The MITFhigh/PD-L1high cells were also enriched for the Tsoi neural-crest like gene set compared to the MITFhigh/PD-L1low cells, which themselves showed a melanocytic transcriptomic profile (Supplementary Fig. S12C). Lastly, the PD-L1high groups among both MITFhigh and MITFlow cells shared the “STTTCRNTTT_IRF_Q6” gene set as the top upregulated transcription factor target gene set compared to their PD-L1low counterparts (Supplementary Fig. S12D). This is an IRF1 target gene set, consisting of genes containing at least one occurrence of the STTTCRNTTT motif in proximity of their transcription start site [49]. Together, this indicates that whilst PD-L1 expression varies both among MITFlow and MITFhigh cells, high IFNγ-induced PD-L1 expression is generally associated with enhanced inflammatory gene expression (Fig. 10B, Supplementary Fig. S12B). Furthermore, among the MITFhigh cells specifically, high PD-L1 expression associates with distinct sub-phenotypes that possess features of dedifferentiation as indicated by a more neural-crest like transcriptional profile compared to MITFhigh/PD-L1low cells (Supplementary Fig. S12C).
留言 (0)