
記住我
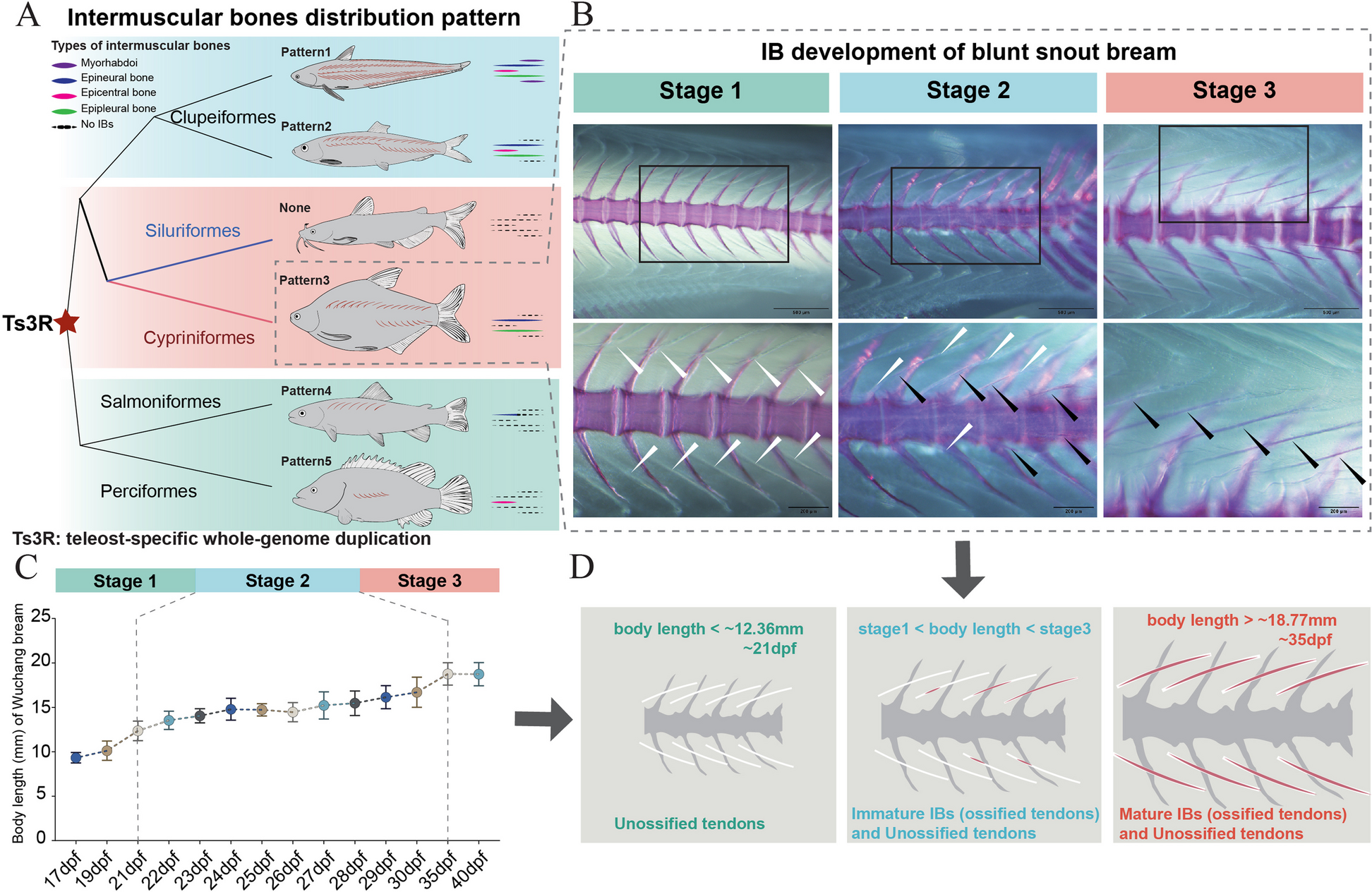




Based on information provided by Patterson and Johnson [14], Yang et al. [20], and Nelson [24], we summarized 5 IB distribution patterns in teleosts (Fig. 1A). In Clupeiformes, there are 2 patterns including four-categories-IB model (epineural bones—ENBs, epipleural bones—EPBs, epicentral bones—ECBs, and myorhabdois—MBs) and three-categories-IB model (ENBs, EPBs, and ECBs); there is no IBs in Siluriformes; Cypriniform fishes possess 2 categories of IBs, ENBs, and EPBs; Salmoniform fishes have only ENBs; and Perciform fishes have only ECBs. IBs are diverse in distribution and found only in teleost fishes [14], which may be attributed to teleost-specific whole-genome duplication (Ts3R).
Fig. 1
The developmental and evolutionary characteristics of IBs (tendon ossification in myosepta). A IB distribution patterns in teleosts. Pattern 1 (Clupeiformes, Japanese grenadier anchovy (Coilia nasus)), epineural bones (ENBs), epipleural bones (EPBs), epicentral bones (ECBs), and myorhabdois (MBs); pattern 2 (Clupeiformes, Atlantic herring (Clupea harengus)), ENBs, EPBs, ECBs; pattern 3 (Cypriniformes, blunt snout bream (M. amblycephala)): ENBs and EPBs; none (Siluriformes, channel catfish (Ictalurus punctatus)), no IBs; pattern 4 (Salmoniformes, rainbow trout (Oncorhynchus mykiss)): ENBs; pattern 5 (Perciformes, Mandarin fish (Siniperca chuatsi)): ECBs. B–D Three developmental stages of IBs in blunt snout bream. B Histological analysis of IB development. White arrows marker tendons in myosepta; black arrows marker ossified tendons (IBs) in myosepta. C The relationship between IB development and body length. D Stage 1: only unossified tendon distribution without IB development (body length < ~ 12.36 mm); stage 2: distribution of both immature IBs (ossified tendons) and unossified tendons (body length ~ 12.36– ~ 18.77 mm); stage 3: distribution of mature IBs (ossified tendons) (body length > 18.77 mm)
Cypriniformes, constituting the largest monophyletic group of freshwater fishes on Earth [20], exhibits the most widespread IB distribution pattern. We focused on the developmental process of IBs in Cypriniformes, using blunt snout bream, a representative species, to observe IB development. We explored the histological structure of IB at different developmental stages (Fig. 1B–D, Additional file 2: Table S1) (stage 1: only unossified tendon distribution without distribution of IB; stage 2: both immature IBs and unossified tendons distribution; stage 3: both mature IBs and unossified tendons distribution) by alizarin red staining. The results are consistent with Chen et al. [6, 17].
Gene expression characteristics of IBsThe evolutionary origin of any new organ typically depends on the recruitment of genes that are originally expressed in other tissues [25]. IBs develop later relative to other tissues and can be considered as a novel tissue. To identify genes recruited in ENBs and EPBs, we compared 13 transcriptomic data from blunt snout bream obtained in this study (Additional file 2: Table S2). In addition, to assess the expression signature of tissue-specific expression genes at three different developmental stages of IBs, we utilized 6 transcriptomic data that were obtained from Liu et al. [26]. Genes specifically expressed in ENBs or EPBs (hereafter referred to as ENB-specific expression genes or EPB-specific expression genes) were defined as those that have a tau index exceeding 0.8 [27] and were expressed most strongly or second most strongly in ENBs or EPBs. One hundred thirty ENB-specific expression genes (Fig. 2A, Additional file 2: Table S3) and 224 EPB-specific expression genes (Fig. 2B, Additional file 2: Table S4) were identified. The main sources of ENB were EPB (70.00%), neural spines (6.92%), operculum (6.92%), suboperculum (3.85%), and vertebra (3.85%) (Fig. 2A); the main sources of EPB were ENB (40.62%), skin (14.73%), rib (10.27%), operculum (9.38%), and neural spines (6.70%) (Fig. 2B). Importantly, 91 IB-specific expression genes by tau were shared by both ENBs and EPBs and enriched in biological processes related to IB development (Fig. 2C, Additional file 2: Table S5, Additional file 2: Table S6). To elucidate the temporal expression characteristics of core genes, time-series analysis was performed. The results of the time series analysis revealed that most core genes exhibited high expression levels during the early stages of IB formation (stage 1 and stage 2), indicating that the design of IB formation occurred during the early developmental stages of blunt snout bream (Additional file 1: Fig. S1). To examine the impact of Ts3R on IB development, we screened for genes undergoing Ts3R in blunt snout bream genome and found 8 IB-specific expression genes by tau (dbx1b, fgf8b, LOC125252216, LOC125264141, LOC125278619, plg, slc16a1a, tfa) undergoing Ts3R (Fig. 2D, Additional file 2: Table S7). These genes were all highly expressed in the early stages of IB development. plg was enriched in blood circulation (GO:0008015 p value = 8.418093e − 04) and extracellular matrix organization (GO:0030198 p value = 2.142292e − 03). tfa was enriched in positive regulation of bone remodeling (GO:0046852 p value = 6.775198e − 04), positive regulation of MAPK cascade (GO:0043410 p value = 2.380401e − 03), and regulation of bone mineralization (GO:0030500 p value = 7.032019e − 03) (Fig. 2D). In summary, ENB-specific expression genes and EPB-specific expression genes were recruited from similar tissues and shared 91 same genes, suggesting they have similar gene expression characteristics and tissue origins. Subsequent analysis focused on the genes shared by ENBs and EPBs. Meanwhile, Ts3R may have an impact on the occurrence of IBs in teleosts (Fig. 1A).
Fig. 2
Gene recruitment and cellular origin of ENB and EPB in blunt snout bream. A Genes recruited to ENBs mainly originated from epipleural bones (EPBs), neural spines, operculum, suboperculum, and vertebra. The expression levels of ENB-specific expression genes were depicted in heatmap. B Genes recruited to EPBs mainly originated from epineural bones (ENBs), skin, rib, operculum, and neural spines. The expression levels of EPB-specific expression genes were depicted in heatmap. C Gene Ontology (GO) analysis of the intersection of ENB-specific expression genes and EPB-specific expression genes. D Temporal expression and GO analysis of genes influenced by Ts3R
Identification of genes related to tendon ossification across speciesThe primitive teleostean condition with regard to intermuscular bones is to have ENBs alone. Epipleural bones are absent in the most primitive teleosts [14]. However, both ENBs and EPBs are present in Cypriniformes but absent in Siluriformes. In Cypriniformes, Nie et al. [10] found that stem cells in myosepta differentiated into osteoblasts and tenocytes and revealed the role of osteoblasts in IB formation. Therefore, we proposed that myoseptal stem cells in IB-containing fish have two differentiation pathways: from stem cell to osteoblast and from stem cell to tenocyte. In contrast, myoseptal stem cells in IB-lacking fish could only differentiate into tenocytes. With a goal of identifying the genes responsible for the formation of IB in the evolution of teleosts, we conducted a comparative transcriptomic analysis of myoseptal tissues in fish with and without IBs. IB-containing group included blunt snout bream and grass carp (Ctenopharyngodon idella); IB-lacking group included yellow catfish (Tachysurus fulvidraco) and channel catfish (Ictalurus punctatus).
For cross-species differential expression analysis, 19,997 one-to-one orthologs in IB-containing group (Additional file 2: Table S8) and 15,818 one-to-one orthologs in four species (Additional file 2: Table S9) were identified. Furthermore, 3302 specific genes in IB-containing group were identified (Fig. 3A, Additional file 2: Table S10). We constructed comparative groups: ENB vs. epineural muscles (ENMs include muscles and epineural tendons) and EPB vs. epipleural muscles (EPMs include muscles and epipleural tendons) within IB-containing group. Differential expressed analysis using GFOLD (v1.1.4) [28] revealed an intersection of 623 IB-containing fish genes by DEA (differential expressed analysis within IB-containing group, DEA) with the same expression trend (Fig. 3B, Additional file 2: Table S11). These genes were enriched in upregulated biological processes (extracellular matrix organization (GO:0030198), ossification (GO:0001503), regulation of angiogenesis (GO:0045765), connective tissue development (GO:0061448), and others) and downregulated biological processes (protein ubiquitination (GO:0016567) and others) (Fig. 3B, Additional file 2: Table S12). Gene set enrichment analysis (GSEA) indicated that upregulated biological processes may be associated with promoting IB development, while downregulated biological processes may inhibit it. We performed similar differential expressed analysis using GFOLD and SCBN (v1.21.0) [29] between IB-containing group and IB-lacking group (details in “Methods”). Ultimately, 209 IB-CDEA genes (cross-species differential expressed analysis between IB-containing group and IB-lacking group, CDEA) with the same expression trend were identified in IB-containing vs. IB-lacking group (Fig. 3C, Additional file 2: Table S13). We divided the two gene sets (623 IB-containing fish genes by DEA and 209 IB-CDEA genes) into 4 categories, including 69 specific genes in IB-containing fish, 79 intersection genes shared by two gene sets, 475 other genes in 623 IB-containing fish genes by DEA, and 130 other genes in 209 IB-CDEA genes, based on homology and gene expression trends (Fig. 3C). In the IB-containing fish group, 11.08% of 623 IB-containing fish genes by DEA were IB-containing-fish specific genes; 12.68% of 623 IB-containing fish genes by DEA were shared by IB-containing group and IB-containing vs. IB-lacking group; 76.24% of 623 IB-containing fish genes by DEA were no significant differences in IB-containing vs. IB-lacking group or did not belong to one-to-one orthologous gene pairs. In IB-containing vs. IB-lacking group, 37.80% of 209 IB-CDEA genes were shared by IB-containing group and IB-containing vs. IB-lacking group; 62.20% of 209 IB-CDEA genes were no significant differences in IB-containing fish group (Fig. 3C). These four categories of gene sets exhibited two different expression trends during the formation of IBs in three developmental stages: one highly expressed in early IB development (Cluster2 and Cluster3) and the other in late IB development (Cluster1) (Fig. 3D).
Fig. 3
Comparative transcriptomic analysis for identifying key genes involved in IB development. A Identification of one-to-one orthologous gene pairs across different species. A total of 19,997 one-to-one orthologous gene pairs were identified in the IB-containing group, and 15,818 one-to-one orthologous gene pairs were identified in the IB-containing vs. IB-lacking fish group. A total of 3302 genes were specific to the IB-containing fish group. B Identification of 623 IB-related differentially expressed genes in the IB-containing fish group. Gene set enrichment analysis was performed on this gene set. C Grouping of differentially expressed genes identified from the IB-containing fish group (left plot) and the IB-containing vs. IB-lacking fish group (right plot). D Expression patterns of genes from different groups in C during IB development. E Lollipop plot displaying IB-containing-fish specific differentially expressed genes and orthologous differentially expressed genes. F Functional network plot of selected differentially expressed genes
The 28 upregulated and 41 downregulated specific expression genes in IB-containing fish were shown in Fig. 3E. clec3bb, cltcl1, six4a, fgfr1b, fgf13a, chrna1, and LOC125277479 were enriched in upregulated biological processes function to IB formation (Fig. 3F, Additional file 2: Table S14), while amfrb was a downregulated specific gene involved in protein ubiquitination in IB-containing fish (Additional file 2: Table S14). Additionally, 37 upregulated and 37 downregulated intersection genes in IB-containing vs. IB-lacking group were also shown in Fig. 3E (tnnt3b, rtn4a, fxr1, zgc:91,999, and pfkfb3 were downregulated in IB-containing fish group but upregulated in IB-containing vs. IB-lacking group). loxa, timp2a, col5a1, lum, thbs2a, mmp2, ryr1b, aspn, tnfrsf11a, and mef2aa were involved in upregulated biological processes functioning to IB formation (Fig. 3F, Additional file 2: Table S14). In short, significant changes in gene expression related to IB formation occurred during the evolution of teleosts. Only a subset of core genes exhibited consistent expression trends, potentially associated with the differentiation of myoseptal stem cells into osteoblasts.
Candidate genes show altered expression in IB-lacking zebrafishThrough comparative analysis of multiple species and tissues, we identified 79 differentially expressed genes that were conserved across species (IB-containing fish (blunt snout bream and grass carp) and IB-lacking fish (channel catfish and yellow catfish)) (Fig. 3E right), 69 specific expression genes in IB-containing fish (Fig. 3E left), and 91 IB-specific expression genes by tau (Additional file 1: Fig. S1C). To screen for genes associated with tendon ossification (Additional file 1: Fig. S2), we further investigated the expression of the above candidate genes in runx2b−/− zebrafish (IB-lacking zebrafish by genome editing, Nie et al. [10]). Upon reanalysis of the transcriptome data (runx2b−/− and runx2b+/+ tail tissues) from Xiao et al. [11] (Additional file 1: Fig. S2), a total of 1840 differentially expressed genes were identified (Additional file 1: Fig. S3, Additional file 2: Table S15). Notably, four specific genes in IB-containing fish (clec3bb, myl2b, actn3b, and ptx3b) (Fig. 4A–C), four differentially expressed genes conserved across four species (krt18b, si:dkey − 282h22.5, si:ch73 − 86n18.1, and si:ch211 − 243a20.3) (Fig. 4D–F), and two IB-specific expression genes by tau (prokr1a and cp) (Fig. 4G–I) exhibited opposite expression patterns in the genome-edited zebrafish group through intersection analysis.
Fig. 4
Comparative analysis of gene expression patterns in runx2b+/+ and runx2b−/− zebrafish. A–C Intersection genes between 69 specific genes in IB-containing fish (Fig. 3E left) and genome editing zebrafish DEGs (differential expressed genes). Expression patterns of intersecting genes in different groups. D–F Intersection genes between 79 differentially expressed genes conserved across species (Fig. 3E right) and genome editing zebrafish DEGs. Expression patterns of intersecting genes in different groups. G–I Intersection genes between 91 IB-specific expression genes by tau and genome editing zebrafish DEGs. Expression patterns of intersecting genes in different groups. Cid, grass carp; Mam, blunt snout bream; Ipu, channel catfish; Tfu, yellow catfish
clec3bb shows specific and elevated expression in tenocytes of zebrafish without IBsIn order to elucidate the single-cell expression atlas of genes associated with IB formation, we reanalyzed the single-cell transcriptome data from runx2b+/+ and runx2b−/− zebrafish tail tissues [10]. The uniform manifold approximation and projection (UMAP) analysis of the scRNA-seq data identified 14 distinct cell clusters (Additional file 1: Fig. S4, Additional file 2: Table S16). To explore the potential differentiation relationship between tenocyte/fibroblast and osteoblast, we investigated the diversity tenocytes, fibroblasts, and osteoblasts in more detail (Additional file 1: Fig. S4, Additional file 2: Table S16). Sub-clustering analysis revealed an unexpectedly large degree of diversity, with 10 transcriptionally distinct clusters (Fig. 5A, Additional file 2: Table S17). There were significant differences in the proportion of most clusters between runx2b+/+ and runx2b−/− zebrafish. The proportions of clec3bb + tenocyte, fibroblast2, and osteoblast1/2 were higher in runx2b+/+ zebrafish than runx2b−/− zebrafish. Conversely, the proportions of tenocyte2, fibroblast1, and mstnb + tenocyte were higher in runx2b−/− zebrafish than runx2b+/+ zebrafish. There were no significant differences between runx2b+/+ and runx2b−/− zebrafish in tenocyte1, myoseptal stem cell, and thbs4b + tenocyte cluster (Fig. 5B). The tenocyte supercluster consisted of four distinct subpopulations, including differentiating tenocyte clusters (tenocyte1 and tenocyte2, which were marked by scxa), clec3bb + tenocyte, thbs4b + tenocyte, and mstnb + tenocyte (Fig. 5B). Clec3b gene is associated with bone mineralization, and its deficiency leads to skull defects in mice [30]. thbs4b controls matrix assembly during development and repair of myotendinous junctions in zebrafish [31]. Spontaneous and targeted mstn loss-of-function mutations lead to double muscle phenotypes in zebrafish [32]. mstnb + tenocyte may be located in the myotendinous junction, serving as the boundary between muscle and tendon. There were 3 sub-clusters in fibroblast supercluster, including fibroblast1 (nt5e), myoseptal stem cell (cxcl12a), and fibroblast2 (fn1a) (Fig. 5B). Cxcl12 is a marker commonly used for hematopoietic stem cells [33], osteoblasts [34], and mesenchymal stem cells of heterotopic ossification [9]. Cells in the myoseptal stem cell cluster may contribute to tendon ossification. Osteoblast supercluster included osteoblast1 and osteoblast2, marked by runx2b. cd81b, a biomarker of exosomes [35], was specific expressed in osteoblast2, suggesting that osteoblasts2 interact with other cells via exosomes.
Fig. 5
The cellular differentiation process of tendon ossification in runx2b+/+ (with IBs) and runx2b−/− (without IBs) zebrafish. A Reclustering of superclusters associated with tendon ossification in runx2b+/+ and runx2b−/− zebrafish. B Proportional analysis and marker gene annotation of the 10 sub-clusters. C RNA velocity analysis of 10 sub-clusters in runx2b+/+ zebrafish. D RNA velocity analysis of 10 sub-clusters in runx2b−/− zebrafish. E Violin plots of candidate genes in Fig. 4
With the hope of uncovering the effect of runx2b deficiency on cell differentiation, RNA velocity analysis was performed in runx2b+/+ and runx2b−/− single-cell data. Myoseptal stem cell could differentiate into osteoblast, tenocyte, and fibroblast in runx2b+/+ zebrafish (Fig. 5C). Due to runx2b deficiency, cell differentiation trajectory was affected in runx2b−/− zebrafish. The ability of myoseptal stem cell to differentiate into osteoblast1/2 was weakened, while the ability to differentiate into fibroblast1/2 and clec3bb + tenocyte was enhanced (Fig. 5D).
The expression of candidate genes identified in the previous sections was also altered in runx2b−/− zebrafish scRNA data, consistent with the bulk RNA-seq (Fig. 4). clec3bb, involved in ossification (GO:0001503), was upregulated in clec3bb + tenocyte of runx2b−/− zebrafish. fgfr1b, involved in connective tissue development (GO:0061448), was upregulated in osteoblast1 and osteoblast2 of runx2b−/− zebrafish. cnn2 was upregulated in all clusters except osteoblast2 in runx2b−/− zebrafish. col2a1b was downregulated in all clusters in runx2b−/− zebrafish. lum, participating in extracellular matrix organization (GO:0030198) and connective tissue development, was downregulated in myoseptal stem cell and osteoblast supercluster of runx2b−/− zebrafish. thbs4b, involved in connective tissue development, was downregulated in fibroblast2 and osteoblast1 of runx2b−/− zebrafish. aspn, involved in ossification, was downregulated in fibroblast2 and osteoblast2 of runx2b−/− zebrafish. loxa, involved in extracellular matrix organization, was upregulated in all clusters except fibroblast1, fibroblast2, and osteoblast2 of runx2b−/− zebrafish. We further validated the expression of other candidate genes (fgfr1b, cnn2, col2a1b, lum, thbs4a, aspn, and loxa) in two IB-development stages of runx2b−/− zebrafish and runx2b+/+ zebrafish, which was consistent with the mRNA expression of clec3bb (higher expression in 60 dpf runx2b−/− zebrafish than in 60 dpf runx2b+/+ zebrafish) (Additional file 1: Fig. S5).
Gene structure and expression analysis of clec3ba and clec3bbClec3b belongs to tetranectin gene family [36], which includes four genes (clec3ba, clec3bb, clec3a, and clec11a) in zebrafish. To explore IB-specific genetic changes of the tetranectin gene family among teleosts, we compared conserved motifs, gene structures, and protein structures from four IB-containing fishes and three IB-lacking fishes. Osteolectin (also known as CLEC11A) is an osteogenic factor that promotes the maintenance of the adult skeleton by promoting the differentiation of LepR + cells into osteoblasts [37]. Clec11a-deficient mice exhibit accelerated bone loss during aging, reduced bone strength, and delayed fracture healing [38]. Motif10 was specific to IB-lacking fish (Additional file 1: Fig. S6). CLEC3A is identified as cartilage-specific antimicrobial peptide in septic arthritis and contributes to cell proliferation through HIF1α signaling pathway [39]. Motif7 was specific to IB-lacking fish (Additional file 1: Fig. S6). C-type lectin domain family 3 member B (CLEC3B), known as tetranectin (TN), is detected in the cytoplasm, extracellular matrix (ECM), and exosomes [40]. Teleosts possess clec3ba and clec3bb under the action of Ts3R, but clec3bb was lost in IB-lacking fish. There were no differences in CLEC3BA conserved motifs between IB-lacking and IB-containing fish. Otherwise, clec3ba genes of IB-containing fish had longer UTR sequences compared to those of IB-lacking fish (Additional file 1: Fig. S6). We further compared the differences in amino acid sequences between CLEC3BA and CLEC3BB in IB-lacking fish and IB-containing fish (Fig. 6A). There were four IB-specific amino acid changes (K26T, N35H, Q60H, and H158R) and a missing fragment of IB-containing fish in CLEC3BA. The presence of conserved amino acids (such as A61, K84, and W152) also implied the conserved nature of gene function between CLEC3BA and CLEC3BB (Fig. 6A).
Fig. 6
IB-specific genetic changes and gene expression features in tetranectin gene family. A Amino acid changes in CLEC3BA and CLEC3BB across IB-containing fishes (Clupea harengus, Danio rerio, Megalobrama amblycephala, Ctenopharyngodon idella) and IB-lacking fishes (Silurus meridionalis, Ictalurus punctatus, Tachysurus fulvidraco). B clec3bb expression in 13 tissues of blunt snout bream. C The expression pattern of clec3bb gene in 24 hpf (hours post fertilization) and 48 hpf zebrafish embryos. D The expression pattern of clec3bb gene in 60 dpf zebrafish by STARmap FISH
Furthermore, clec3bb, as an IB-containing fish specific gene, was expressed more highly in ENBs and EPBs than in any other tissues (Fig. 6B). It was also expressed in myoseptas of zebrafish embryo (24 and 48 hpf, hours post fertilization) (Fig. 6C) and in ossified tendons (IBs) of adult zebrafish (Fig. 6D).
clec3bb promotes osteoblast2 mineralization in myosepta of IB-containing fishC-type lectin domain family 3 member B (CLEC3B) is a transmembrane Ca2+-binding protein located in cell plasma, extracellular matrix, and exosomes [41]. In runx2b+/+ zebrafish, clec3bb was specifically expressed in clec3bb + tenocytes, and its expression products functioned to osteoblast2 through exosomes to promote osteoblast2 mineralization (Fig. 7A). Osteoblast2 specifically expressed exosome marker cd81b (Fig. 5B), suggesting that osteoblast2 may interact with exosomes. In runx2b−/− zebrafish, runx2b deficiency inhibited the formation of osteoblast1 and osteoblast2, leading to the loss of exosomal-clec3bb-targeted cells and resulting in redundant expression of the clec3bb gene (Fig. 7B). Slingshot analysis revealed that runx2b deficiency resulted in differentiation from thbs4b + tenocytes to clec3bb + tenocytes (Fig. 7A–B). We selected biomarkers of exosome (cd81a, cd81b, cd9a, cd9b, tsg101, and cd63) for qPCR analysis in two IB-development stages of runx2b−/− zebrafish (without tendon ossification) and runx2b+/+ zebrafish (with tendon ossification). The results showed that some exosome biomarkers (cd9a, cd9b, tsg101, and cd63) were expressed at higher levels in runx2b−/− zebrafish than runx2b+/+ zebrafish, while cd81b (an osteoblast2 specific marker) and cd81a (paralogous gene of cd81b) were elevated in 60 dpf runx2b−/− zebrafish. The expression changes of cd81b and cd81a suggested that these genes were related to osteoblast development, particularly cd81b, which was consistent with the mRNA expression of clec3bb (Fig. 7C–D). The results of FISH showed that the three gene expression sites (sp7, cd81b, clec3bb) were similar around IB with a resolution of 200 nm (Fig. 7E). In brief, these results suggested that clec3bb may promote osteoblast2 mineralization via osteoblast-specific exosomes.
Fig. 7
Graphic summary of the role of clec3bb in tendon ossification. A We discovered that myoseptal stem cell could differentiate into osteoblasts in wild type zebrafish by Slingshot analysis. Osteoblast2 further mineralized and formed IBs. Myoseptal stem cell could also differentiate into clec3bb + tenocyte, ultimately forming thbs4b + tenocyte. clec3bb showed specific expression in clec3bb + tenocytes. B After runx2b deficiency, the process of myoseptal stem cell differentiating into osteoblasts was inhibited. The process of differentiation from myoseptal stem cell to clec3bb + tenocytes undergone alterations. clec3bb expression level was increased in clec3bb + tenocytes of runx2b−/− zebrafish. C mRNA relative expression of biomarkers of exosome (cd81a, cd81b, cd9a, cd9b, tsg101, and cd63) in 15 dpf (stage without IBs/tendon ossification in wild type zebrafish) of runx2b−/− zebrafish and runx2b+/+ zebrafish and in 60 dpf (stage with IBs/tendon ossification in wild type zebrafish). D Comparison of gene expression between runx2b−/− and runx2b+/+ zebrafish (t-test, two-sided, *** (p value < 0.001)). Error bars represent
留言 (0)