
記住我


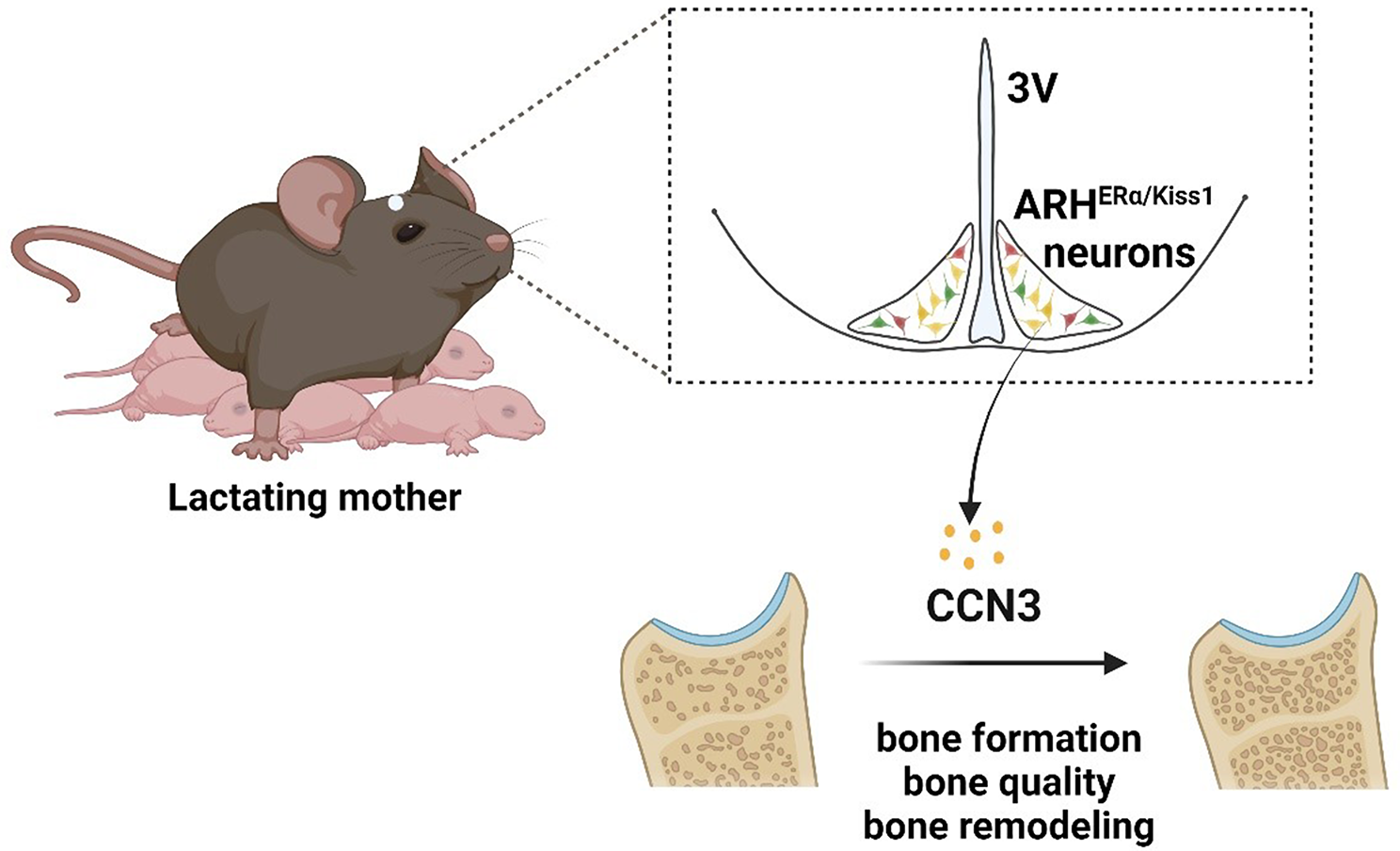




All patients presented with primary infertility of undetermined etiology, having undergone several unsuccessful in vitro fertilization-embryo transfer (IVF-ET) cycles. In family 1, the individual II-1 endured eight years of primary infertility and underwent three oocyte retrieval attempts. Unfortunately, all 65 oocytes retrieved were unsuitable for IVF or ICSI due to immaturity (GV or MI stages) or morphological abnormalities (Fig. 2A), as detailed in Table 1. Notably, during the third retrieval, in vitro maturation (IVM) was performed for nine MI-stage oocytes, resulting in four oocytes exhibiting two-cell-like morphology, suggestive of large polar bodies (Fig. 2A).
Table 1 Oocyte characteristics of IVF and ICSI attempts for the affected individualsIn family 2, proband II-1, after five years of diagnosed primary infertility, underwent two ICSI attempts. From 31 retrieved oocytes, only 14 were mature (MII stage), while four remained immature and the rest were abnormal. This cycle resulted in three 3PN zygotes, which consistently arrested at the 2- to 5-cell stage, as shown in Table 1; Fig. 2A.
In family 3, patient II-1 underwent a single IVF cycle, yielding two 2PN zygotes and one 0PN zygote; however, all embryos arrested at 2- to 4-cell stage. Subsequent attempts yielded only three immature or abnormal oocytes (Table 1).
Family 4’s patient II-1 displayed a higher proportion of mature oocytes (27 MII out of 39 retrieved), yet the remaining oocytes were arrested at either the GV or MI stage. Of the nine zygotes obtained, all embryos ceased development at the 2- to 5-cell stage (Table 1; Fig. 2A).
Lastly, in family 5, patient II-1 from a consanguineous background underwent two IVF/ICSI cycles, retrieving 26 oocytes. Of these, 9 were arrested at the GV stage, 2 at the MI stage, with the remainder exhibiting morphological abnormalities or degeneration (Fig. 2A). Detailed clinical characteristics of the retrieved oocytes are summarized in Table 1.
Identification of novel PATL2 variants in oocyte maturation defect patientsWhole-exome sequencing (WES) was employed to identify pathogenic variants in patients exhibiting oocyte maturation defects (OMD). Through this analysis, we identified three novel missense mutations, one novel frameshift mutation, one novel splicing mutation, and one previously reported splicing mutation in the PATL2 gene. In family 1, the affected individual carried biallelic missense mutations: c.1201G > T (p.V401F) and c.1204 C > T (p.R402W), inherited from their father and mother, respectively, as depicted in Fig. 1A. Her two sisters, carrying either heterozygous or wild-type variants of PATL2, exhibited normal fertility. In family 2, the patient harbored a previously reported homozygous splicing mutation c.223 − 14_223-2delCCCTCCTGTTCCA (p.R75Vfs*21), with no available parental inheritance data (Fig. 1B). Family 3’s patient possessed compound heterozygous frameshift mutations: c.1284delA (p.E428Dfs*9) and c.223 − 14_223-2delCCCTCCTGTTCCA (p.R75Vfs*21), inherited from her mother and father, respectively (Fig. 1C). Similarly, the patient in family 4 exhibited compound heterozygous mutations c.1271T > C (p.L424S) and c.223 − 14_223-2del (p.R75Vfs*21), following the parental inheritance pattern (Fig. 1D). The individual from family 5 carried a homozygous splicing mutation c.1613 + 2_1613 + 3insGT with unknown inheritance data (Fig. 1E), verified to induce a frameshift (p.Phe539Cysfs*19) using mini gene assay (Figure S1A-E).
Fig. 1
Identification of PATL2 gene mutations in five unrelated infertile individuals. (A-E) Pedigrees of the 5 families affected by female infertility with Sanger sequencing confirmation below. Squares denote male family members, circles denote female members, solid circles denote affected individuals, and equals signs represent infertility. Red arrows highlight the mutation sites
The pathogenic effects of three novel variants (the missense mutation p.V401F, the frameshift mutation p.E428Dfs*9, and the splicing mutation p.F539Cfs*19) and two newly identified variants (p.R402W and p.L424S) in a recently reported study [38] were not validated. The novel variants are absent from public databases (Table 2), indicating exceedingly low frequency. They are situated within the highly conserved PAT1 domain of PATL2, spanning species from Xenopus laevis to Homo sapiens (Fig. 2B). The frequency of the reported mutation p.R75Vfs*21 was recorded at approximately 7.2 × 10− 6 mutation frequency in the gnomAD exome database (Table 2). Three-dimensional structural analysis of PATL2 suggested that the missense mutations (V401F, p.R402W, and p.L424S) potentially disrupt hydrogen bonding, thereby destabilizing the protein (Fig. 2C). Pathogenic predictions using tools like SIFT and PolyPhen classify these variants as possibly or probably damaging (Table 2). Collectively, these findings underscore the pivotal role of PATL2 mutations as a genetic basis for human OMD.
Table 2 Overview of the PATL2 mutations observed in the five familiesFig. 2
Effects of PATL2 gene mutations on oocyte and embryo development and protein structure in female infertile patients (A) Bright field images showing the developmental stages of oocytes and early embryos from control individuals and patients from Families 1, 4, and 5 with PATL2 mutations. Images include germinal vesicle (GV) oocytes, metaphase I (MI), and metaphase II (MII) oocytes, as well as various stages of embryos, such as the 8-cell stage and blastocyst formation in control, and 2-cell, 4-cell, and 5-cell stages in patients. Scale bars = 20 μm. (B) Schematic representation of the PATL2 protein and the distribution of PATL2 variants (the corresponding amino acid sequences) in PATL2 protein. The conservation of these mutation sites across different species (human, primates, mouse, pig, and Xenopus) with mutated residue marked in yellow. (C) PATL2 missense variants encoding amino acid disrupted the ion pairs formed by wild-type PATL2 protein. The models compare the wild-type and mutant amino acid residues (Val401, Arg402, and Leu424) within the protein structure, illustrating how the mutations (Val401Phe, Arg402Trp, and Leu424Ser) alter the local conformation of the PATL2 protein
Pathogenic effects of PATL2 variants in protein propertyNext, we used HEK293T cells to determine the functional properties of these PATL2 variants compared to wild-type PATL2. We observed that the three PATL2 variants had the similar subcellular localization with wild-type PATL2 in the cytoplasm by immunofluorescence (Fig. 3A). However, these mutations led to decreased PATL2 protein levels, especially those of the p.V401F, p.R402W, and p.E428Dfs*9 variants (Fig. 3B-C). We also microinjected cRNAs of HA-PATL2 or different variants into mouse GV oocytes, the localization was not changed but the protein of PATL2 variants were significantly decreased (Fig. 3D-E).
Fig. 3
Effects of PATL2 gene mutations on protein expression and localization in cells and oocytes. (A) Representative immunofluorescence images showing the expression and localization of FLAG-tagged PATL2 protein (green) in 293T cells. The cells were transfected with vectors expressing wild-type (WT) or mutant PATL2 (V401F, R402W, E428Dfs*9, L424S, and F539Cfs*19). DAPI (blue) was stained for visualization of DNA. Scale bar = 10 μm. (B) Western blot showing the expression levels of FLAG-tagged PATL2 in 293T cells transfected with vectors expressing WT or mutant PATL2 proteins. The blot was probed with anti-FLAG and anti-GAPDH antibodies, with GAPDH serving as a loading control. (C) Bar graph quantifying the intensity of FLAG-tagged PATL2 protein relative to GAPDH from the western blot analysis in (B)). Data are expressed as mean ± SD. Two-tailed Student’s t-test; ***P < 0.001, *P < 0.05, ns: not significant. (D) Representative immunofluorescence images showing the expression and localization of HA-tagged PATL2 protein in oocytes from mice. Fully-grown oocytes (GV stage) were microinjected with cRNAs encoding WT or mutant PATL2 (V401F/R402W, E428Dfs*9, L424S, F539Cfs*19). HA (green) and DAPI (blue) staining indicate the localization of PATL2 and the nuclei, respectively. Scale bar = 10 μm. (E) Bar graph quantifying the intensity of HA-tagged PATL2 protein in oocytes from (D). Data are expressed as mean ± SD. Two-tailed Student’s t-test; ***P < 0.001, *P < 0.05. (F) The bright-field images showing the oocyte stages after 16 h of in vitro maturation in siNC or siPatl2 group. Scale bar = 100 μm. (G-H) The bar graphs showing the GVBD rate (G) and MII rate (H) after siNC or siPatl2 microinjection in GV oocytes. Three independent experiments were performed. Data are expressed as mean ± SD. Two-tailed Student’s t-test; **P < 0.01; ns, no significance. (I) RT-qPCR showing the Patl2 mRNA level in MII oocytes with siNC or siPatl2 siRNA microinjection. Data are expressed as mean ± SD. Statistical significance was determined using unpaired t-test; ***P < 0.001. (J) Immunofluorescence images showing the subcellular localization of PATL2 (red) and FITC-α-Tubulin (green) in MII oocytes undergoing knockdown of Patl2 from GV stage. DAPI was used to stain chromosome (blue). Scale bar = 10 μm. (K) The bright-field images showing the oocytes after microinjection of negative control or mouse Patl2 siRNAs combined with or without wild-type human HA-PATL2 or HA-PATL2 variants cRNAs and culture in medium with 2.5 µM milrinone for 12 h, followed by release to maturation (n > 120 oocytes each group). Scale bar = 100 μm. (L) The bar graphs showing the percentage of GV, MI and MII in K. Data are expressed as mean ± SD
Next, we examined the functional impact of PATL2 mutations on oocyte meiosis. We firstly microinjected siRNAs targeting negative control (siNC) or mouse Patl2 (siPatl2) using mouse fully-grown GV oocytes. In siPatl2 group, the GV breakdown was unaffected; however, the percentage of MII oocytes was slightly decreased compared to the siNC group (Fig. 3F-H). RT-PCR and immunofluorescence analysis confirmed efficient depletion of Patl2 following siRNA treatment (Fig. 3I-J). We then microinjected the HA-PATL2 cRNAs and various mutant cRNAs to assess their ability to rescue oocyte maturation. As expected, wild-type HA-PATL2 successfully rescued the MII rate, whereas the mutant variants exhibited reduced rescuing effects (Fig. 3K-L). Notably, two frameshift variants, HA-PATL2E428Dfs*9 and HA-PATL2F539Cfs*19, completely failed to rescue oocyte maturation compared to siPatl2 group (Fig. 3K-L). These findings indicate that PATL2 mutations primarily result in protein instability and a consequent functional loss in varying degrees.
Impact of PATL2 variants on the protein interactome and cell cycle regulationSince the typical OMD observed in the patient from family 1 harboring two PATL2 variants, we constructed an HA-tagged PATL2V401F/R402W plasmid to assess differences in protein interactions compared to the wild-type PATL2. Using co-immunoprecipitation followed by mass spectrometry (MS/MS), we identified 1221 proteins that exhibited a ten-fold greater interaction intensity with the wild-type PATL2 relative to the vector control (Fig. 4A). This analysis revealed a total of 1019 PATL2V401F/R402W-binding proteins with at least a ten-fold increased interaction intensity relative to the HA vector group (Fig. 4B). Further comparison between the HA-PATL2V401F/R402W mutant and the HA-tagged wild-type PATL2 identified 1305 up-regulated and 950 down-regulated binding proteins (Fig. 4C). Approximately 920 proteins had protein interactions between wild-type or PATL2V401F/R402W mutant (Fig. 4D). Notably, 436 proteins demonstrated decreased interaction intensity in the PATL2V401F/R402W mutant compared to the wild-type PATL2 with at least a 30% reduction in binding intensity (Fig. 4E). These 436 proteins were subjected to gene ontology (GO) analysis, the PATL2V401F/R402W mutation-affected proteins are primarily involved in critical biological processes, such as the cell cycle, regulation of sister chromatid segregation, and regulation of meiotic cell cycle, exemplified by CDC23, MAD2L1, and APC1 (Fig. 4F).
Fig. 4
The altered protein interactome caused by PATL2 mutations associated with oocyte meiotic maturation (A-C) Scatter plots illustrating differential protein interactions in cells expressing FLAG-tagged wild type (WT) PATL2 relative to vector control (Vec) in (A), PATL2V401F/R402W mutant (Mut) relative to Vec (B), and PATL2V401F/R402W mutant relative to WT PATL2 (C). The upregulated (red) and downregulated (green) interacting protein numbers were indicated. Ten-fold was set as a threshold in (A) and (B), and three-fold was used as a threshold in (C). (D) The Venn diagram illustrating the number of shared and unique interacting proteins in WT vs. Vec and Mut vs. Vec comparisons exhibiting a fold change greater than 10 in wild-type versus vector (A) and mutant (B) conditions. (E) Venn diagram showing the overlap of proteins interacting with WT PATL2 but reduced binding intensity in mutant PATL2. (F) Bubble chart displaying biological process terms for downregulated proteins in PATL2 mutant conditions. (G) Representative Western blot images showing the interaction between FLAG-PATL2 (both WT and mutant) and CDC23, MAD2L1 and TUT7. (H) Quantitative analysis of coimmunoprecipitation intensity for CDC23, MAD2L1 and TUT7, compared across FLAG vector, FLAG-PATL2, and mutant conditions, illustrating altered affinity in mutant samples. (I) Representative immunofluorescence images showing the subcellular localization of FLAG-CDC23 (red) co-expressed with HA-PATL2 or PATL2V401F/R402W (green) mutant in GV oocytes. DAPI (blue) indicates the chromosome. Scale bar = 10 μm. (J-K) Immunofluorescence images and quantitative analysis of proximity ligation assay (PLA) signals in GV oocytes co-expressing FLAG-CDC23 with either HA-tagged wild-type PATL2 or mutant PATL2V401F/R402W mutant, indicating decreased interaction between HA-PATL2 V401F/R402W mutant and FLAG-CDC23 in oocytes. Scale bar = 10 μm
Co-immunoprecipitation experiments demonstrated that ectopic PATL2 interacts robustly with endogenous proteins such as CDC23 and MAD2L1 in 293T cells (Fig. 4G-H). However, the binding level of the PATL2V401F/R402W mutant with these proteins was significantly reduced (Fig. 4G-H). We further verified the interaction in oocyte through microinjecting cRNAs mix of FLAG-CDC23 and HA-PATL2 or HA-PALT2V401F/R402W mutant (Fig. 4I). FLAG-CDC23 and HA-PATL2 (both WT and PALT2V401F/R402W mutant) displayed uniform distribution in both nucleus and cytoplasm (Fig. 4I). Consistently, proximity ligation assays (PLA) showed that significant PLA spots were observed in wild-type HA-PATL2 and FLAG-CDC23, however, the PLA spots number was significantly decreased between HA-PATL2V401F/R402W and FLAG-CDC23 (Fig. 4J-K). These results indicate that PATL2 V401F/R402W variant affects the protein interaction with cell cycle related proteins.
PATL2 binds with CDC23 and stabilizes the CDC23 protein level in cells and mouse oocytesUsing co-immunoprecipitation, we found that ectopic FLAG-PATL2 could interact with endogenous CDC23, MAD2L1 and APC1, whereas it was decreased in PATL2 mutant groups in different extent (Fig. 5A). The interaction between PATL2 and CDC23 or MAD2L1 were not affected with RNase A treatment in co-immunoprecipitation assay (Fig. 5B), indicating their interactions independent of RNAs. Since CDC23 is a key protein for oocyte cell cycle progression and the biallelic mutations in CDC23 cause human OMD and female infertility [17], we determined whether CDC23 is a downstream effector of PATL2 in cell cycle regulation. Ectopic HA-PATL2 and FLAG-CDC23 in 293T cells interacted with each other, and the binding intensity displayed slightly alteration in mutant groups (Fig. 5C). Notably, wild-type PATL2 overexpression increased the protein level of ectopic FLAG-CDC23 protein level, whereas this effect was mild in PATL2-mutant groups (Fig. 5C-D).
Fig. 5
PATL2 interacts with CDC23 and stabilizes its protein level. (A-B) Western blot analysis showing the co-immunoprecipitation (coIP) results of CDC23, MAD2L1, and APC1 using an anti-FLAG antibody in cells expressing FLAG-tagged wild-type (WT) or various mutant forms of PATL2 without RNase A (A) or with 50 ng/ul RNase A treatment (B). (C) Western blot results following CoIP assay showing the interaction of PATL2 and CDC23 in cells expressing HA-tagged PATL2 variants with FLAG-CDC23. (D) Quantification of FLAG-CDC23 intensities normalized to GAPDH in input in (B). Statistical relevance is indicated using one-way ANOVA; *P < 0.05; ***P < 0.001. (E) Immunofluorescence staining of CDC23 (green) in GV oocytes (n > 30) overexpressing HA-tagged WT and mutant PATL2. DAPI was used to stain nuclei (blue). Scale bar = 10 μm. (F) Quantification of fluorescence intensity of CDC23 from (D). Data are expressed as mean ± SD. Statistical significance was determined using a one-way ANOVA; *P < 0.05, **P < 0.01, ***P < 0.001. (G) Immunofluorescence images showing the subcellular localization of and CDC23 (red) and FITC-α-Tubulin (green) in MII oocytes undergoing knockdown of Patl2 from GV stage. DAPI was used to stain chromosome (blue). Scale bar = 10 μm. (H) Western blot analysis of PATL2 and CDC23 on MII oocytes with or without Patl2 knockdown. DDB1 is used as a loading control
Next, we tested whether PATL2 stabilizes CDC23 protein level through protein interaction. We performed CHX chasing assay, and found that PATL2 could stabilize the protein degradation of CDC23, whereas the effect of PATL2 mutants on CDC23 degradation is significantly impaired (Figure S2A-B). Next, we microinjected cRNAs of HA-PATL2 and five variants into mouse GV oocytes and determined the endogenous CDC23 level. We found that PATL2 overexpression in GV oocytes increased the endogenous CDC23 protein level, whereas CDC23 levels were significantly decreased in PATL2 mutant groups (Fig. 5E-F). We also examined the localization and expression of CDC23 in Patl2-knockdown (siPatl2) oocytes. Patl2 knockdown did not change the localization of CDC23 in cytoplasm and at the spindle (Fig. 5G). However, the fluorescent signal of CDC23 was remarkably reduced (Fig. 5G), and immunoblotting confirmed that the protein level of CDC23 was significantly decreased in oocytes with siPatl2 microinjection (Fig. 5H). Taken together the results of PLA in oocytes, these results indicated that PATL2 binds with and stabilizes the CDC23 protein level in oocytes.
PATL2 maintains the protein levels of APC1 and MAD2L1 in mouse oocytesWe also examined the protein levels of other two interacting proteins (APC1 and MAD2L1) of CDC23. As expected, the protein levels of APC1 and MAD2L1 were significantly reduced in fully-grown GV oocytes following Patl2 knockdown (Figure S2C). Immunofluorescence analysis further confirmed that Patl2 knockdown led to a decrease in protein intensity of both MAD2L1 and APC1 (Figure S2D-G). These results indicate that PATL2 is crucial for the maintenance of APC1 and MAD2L1 protein levels in oocytes.
Human PATL2 governs maternal mRNA homeostasis during human oocyte maturationPrevious study has reported that Patl2 loss affects RNA homeostasis in mouse oocyte [31]. Our co-IP and MS/MS results showed that PATL2 interacts with TUT7 and TUT4 (Fig. 4G and Table S4), two terminal uridylyltransferases with redundancy in regulation of RNA degradation. To determine whether PATL2 also regulates mRNA homeostasis in human oocytes, we performed low-input RNA-seq using GV, MII oocytes and day 3 early embryos from unidentified control patients (referred as WT group in Fig. 6). These samples include one GV, one MII, and five arrested embryos from the patient in family 3 with PATL2L424S/R75Vfs*21, and two GV and two MII oocytes with large polar body from the patient from family 5 with PATL2F539Cfs*19 (Fig. 6A and Figure S3A). Using PCA analysis, the transcriptome of PATL2 mutant GV oocytes or day 3 early embryos were significantly different from those of control (Ctrl) patients (Fig. 6B), whereas the distribution of MII oocytes between control and PATL2-mutant groups were comparable (Fig. 6B). The mutation of PATL2 hardly affects the PATL2’s mRNA level in GV oocytes but it was upregulated in MII and early embryos at day 3 (Figure S3B). After calibration with ERCC spike-in, we firstly compared the total mRNA level using TPM (Transcripts Per Kilobase per Million mapped reads) between control and PATL2 mutant samples. In control group, the total mRNA levels were gradually decreased from GV oocytes, MII oocytes to day 3 early embryos (Fig. 6C). In contrast, the mRNA level was significantly decreased in PATL2-mutant oocytes at GV stage (Fig. 6C). However, the total mRNA levels of MII oocytes or day 3 early embryos were comparable between control and PATL2 mutant groups (Fig. 6C). We analyzed the expression distribution of detected transcripts, and found that the gene counts with TPM over 10 in PATL2-mutant GV oocytes is much less than those in control group (Fig. 6D), which indicate that the less mRNA dosage in PATL2-mutant GV oocytes is due to the low mRNA expression of highly-expressed genes. In contrast, in MII stage or day 3 embryos, the high-expression genes (TPM > 10) number in PATL2-mutant group were higher than control group (Figure S3C).
Fig. 6
Transcriptomic profiling of oocytes and early-stage embryos harboring PATL2 mutations. (A) Experimental design diagram depicting the groups studied: control (Ctrl) and PATL2 mutant (Mut), including the specific mutations PATL2L424S/R75Vfs*21 and PATL2F253Cfs*19. Samples were collected for RNA-seq including germinal vesicle (GV) oocytes, metaphase II (MII) oocytes, and day 3 embryos. (B) Principal component analysis (PCA) illustrating transcriptomic segregation between control and PATL2-mutant samples, labeled by developmental stage. (C) Graphical representation of normalized total RNA reads using TPM across developmental stages between control and PATL2-mutant samples. Two-tailed Student’s t-test; *P < 0.05, ns: no significance. (D) Bar graph showing the mean transcript counts based on the gene expression level. (E) Volcano plots detailing differential expression genes across the GV, MII, and day 3 embryo stages between control and PATL2-mutant samples. The upregulated (red) and downregulated (blue) gene numbers were indicated. (F) Sankey diagram representing changes in expression status across the GV, MII, and day 3 embryo stages between control and PATL2-mutant samples, highlighting the continuity and shift between expression categories. (G) Quadrant diagram delineating the shared gene counts between MII/GV stages and PATL2 mutants relative to controls using log2 fold change in TPM values over 1. (H) Venn diagram showing the distribution and overlap of downregulated transcripts between MII and GV stages under PATL2 mutant conditions, as compared to controls. (I) Bar chart showing enriched Gene Ontology (GO) terms of the downregulated genes both in GV and MII oocytes with PATL2 mutation. (J) KEGG pathways enrichment of down-regulated genes at various developmental stages. (K) Heatmaps showing expression patterns of cell cycle related genes across the GV, MII, and day 3 embryo stages between control and PATL2-mutant samples, colored by relative expression levels from low (blue) to high (red)
Detailed comparative analysis supported an abnormal transcriptome in PATL2-mutant GV oocytes, with significant differences (745 transcripts with reduced abundance, 487 with increased abundance) using a threshold with |log2fold-change| > 1 and padj < 0.05 (Fig. 6E-F). These differences became more pronounced in the MII oocytes (1033 transcripts with reduced abundance, 1170 with increased abundance) and day 3 embryos (1145 transcripts with reduced abundance, 1231 with increased abundance) (Fig. 6E-F). In addition, a quadrant diagram revealed that among 1170 transcripts significantly upregulated in PATL2-mutant MII oocytes, 660 genes should be dramatically degraded across oocyte maturation (Fig. 6G), indicating an obvious delay in mRNA clearance during GV-MII transition with PATL2 mutations (Fig. 6G). We also analyzed the percentage of zygotic genome activation (ZGA) and M-decay genes in down-regulated genes in day 3 embryos. We found 41% (457 in 1145 genes) downregulated genes in PATL2-mutant day 3 embryos were ZGA genes, and 27% (336 in 1231 genes) upregulated genes were M-decay genes (Figure S3D). These results suggested that PATL2 is critical for regulating the mRNA dosage in human immature oocytes and the mRNA degradation in MII oocytes and early embryos.
We also analyzed the conserved down-regulated genes (228 genes) both in GV and MII oocytes (Fig. 6H), which may partially explain the GV arrest. The gene ontology (GO) analysis found that the transcripts relative to RNA polymerase II transcription regulator complex, protein kinase complex and cyclin-dependent protein kinase holoenzyme complex were affected (Fig. 6I). Analysis of functional enrichment data of down-regulated genes with PATL2 mutations at each stage showed that a variety of fundamental biological processes are involved: RNA splicing, mRNA processing and mitochondrial translation at the GV stage (Figure S3E); mRNA processing, chromosome segregation, actin cytoskeleton and spindle organization at the MII stage (Figure S3F); ncRNA processing and ribosome biogenesis in day 3 embryos (Figure S3G). Notably, pathways associated with cell cycle were significantly enriched in down-regulated genes of PATL2-mutant group at the GV, MII stage and day 3 embryos (Fig. 6J-K), and RNA polymerase and RNA degradation was only enriched in day 3 embryos (Fig. 6J). We analyzed the expression level of PATL2-binding proteins relative to oocyte meiosis using the TPM in RNA-seq. All the gene expression (CDC23, ANAPC1, ANAPC2, ANAPC5 and MAD2L1) were comparable between two group at each stage (Figure S3H). However, we noted that the mRNA levels of CCNB1 and CCNE1 were significantly decreased in GV oocytes (Fig. 6K and Figure S3I), and CDC20 was decreased in MII oocytes with PATL2 mutation (Fig. 6K and Figure S3I). WEE2, a maternal gene with mRNA degradation from MII to day 3 embryo, was upregulated in PATL2-mutatnt embryos at day 3 (Fig. 6K and Figure S3I). In summary, these results suggested that PATL2 mutations affected cell cycle-related genes expression and degradation required for human oocyte maturation and early embryonic development.
PATL2 mutation impeded mRNA decay in human oocytes and embryosTo investigate the impact of PATL2 mutations on maternal mRNA degradation in human oocytes and embryos, we analyzed the mRNA dynamics in GV oocytes, MII oocytes, and day 3 embryos using our RNA-seq data. Maternal mRNAs with reliable sequence annotations and a TPM > 2 in the control group at any stage (a total of 38,167 genes) were selected for analysis. Genes were classified into nine categories based on changes in their mRNA levels, defined as a significant 2-fold or greater downregulation or upregulation between stages. Three clusters of degraded maternal mRNAs were identified: Cluster I (1,467 genes), which degraded from GV to MII and remained stable post-fertilization; Cluster II (5,657 genes), which remained stable from GV to MII but degraded in day 3 embryos; and Cluster III (1,671 genes), which exhibited continuous degradation from GV to day 3 embryos (Fig. 7A). In oocytes and embryos with PATL2 mutations, mRNA decay was impaired across all three clusters, suggesting a defect in maternal mRNA degradation.
Fig. 7
PATL2 binds with TUT7 in oocytes and promotes maternal mRNA decay. (A) Degradation patterns of human maternal transcripts at the GV, MII oocytes and day 3 embryos. The green line and red line represent the median expression levels of the cluster in control and PATL2-mutant group, respectively. (B) Representative immunofluorescence images showing the colocalization of ectopic HA-PATL2 (green) and TUT7 (red) in human MII oocytes. The human MII oocytes with or without microinjection with HA-PATL2 cRNAs were stained with anti-HA and anti-TUT7 antibody co-stained with DAPI. Scale bar = 20 μm. (C) Fluorescence intensity of HA-PATL2 and TUT7 of the white line in (B) was measured. (D) The graph showing the intensity of TUT7 in (B). (E) In situ proximity ligation assays (PLA) showing the interaction between HA-PATL2 and TUT7 in human oocytes with or without microinjection of HA-PATL2 cRNAs. Scale bar = 20 μm. (F) The graph showing the PLA spots numbers quantified by Image J software. (G) The western blot results showing the coimmunoprecipitation of ectopic expression of HA-PATL2 and PATL2 variants in 293T cells. The anti-HA and anti-TUT7 were immunoblotted. (H) Bar graph showing expression levels of TUT7 in RNA-seq of PATL2-mutant and control GV, MII, and Day 3 embryos. (I) RT-qPCR results showing the Tut7 mRNA level in MII oocytes with siNC and siPatl2 knockdown (n = 3 biological replicates). (J) Representative images showing TUT7 (red) in MII oocytes microinjected with siNC and siPatl2 at GV oocyte. The spindles were stained with FITC-α-Tubulin (green) and chromosomes were stained with DAPI (DAPI). Scale bar = 20 μm. (K) Quantifications of TUT7 intensity in (I). An average fluorescence intensity is measured by single oocyte and is plotted as a single dot. Data in (D), (F), (H), (I) and (K) are expressed as mean ± SD. Two-tailed Student’s t-test; ***P < 0.001; ns, no significance
The remaining clusters included: Cluster IV (2,105 genes), stable across all stages; Cluster V (231 genes), which elevated from GV to MII and remained stable from MII to day 3 embryos; Cluster VI (2,656 genes), stable from GV to MII but elevated from MII to day 3 embryos; Cluster VII (244 genes), consistently elevated from GV to the embryonic stage; Cluster VIII (1,540 genes), which degraded from GV to MII and elevated post-fertilization; and Cluster IX (892 genes), which elevated from GV to MII but degraded after fertilization (Figure S4A). Notably, the mRNAs transcribed during zygotic genome activation (ZGA), identified in Cluster VI, were impaired in day 3 embryos with maternal PATL2 mutations. These results demonstrate that PATL2 mutations disrupt both mRNA decay and ZGA (Fig. 8).
PATL2 interacts with CPEB1 and TUT7 in human oocytes and embryosGiven the evidence that PATL2’s interaction with CPEB1 to modulate RNA homeostasis in mouse oocytes [31] and interaction with TUT7 in Fig. 4G, we extended these findings to human oocytes and embryos. We investigated the roles of PATL2 in mRNA accumulation and decay by examining its interactions with CPEB1 and TUT7, respectively. Through extraction of the TPM of CPEB1 mRNAs in our RNA-seq data (Table S5), we found CPEB1 mRNAs are comparable between control and PATL2-mutatnt oocytes, but was upregulated in day 3 embryos (Figure S4B), indicating degradation defect. Following microinjection of HA-tagged PATL2 into human oocytes or 3PN zygotes, we observed colocalization of HA with both CPEB1 (Figure S4C). Proximity ligation assays (PLA) using antibodies against HA and CPEB1 revealed a higher number of PLA signals in the HA-PATL2 group compared to controls (Figure S4D-E). Similar to human oocytes, the mRNA level of Cpeb1 in mouse MII oocytes with Patl2 knockdown was unaffected (Figure S4F). These suggest a conserved role of PATL2 and CPEB1 in maintaining mRNA accumulation across mouse and human oocytes.
Similarly, colocalization of HA-PATL2 and endogenous TUT7 was observed in human oocytes (Fig. 7B-C). Notably, HA-PATL2 overexpression induced a protein increase of TUT7 (Fig. 7D). The enhanced PLA signals were noted between HA-PATL2 and TUT7 in oocytes relative untreated control oocytes (Fig. 7E-F). Using coimmunoprecipitation, the wild-type PATL2 binds with TUT7, and the binding intensities between PATL2 mutants (except to PATL2F539Cfs*19) and endogenous TUT7 were decreased (Fig. 7G). We then evaluated the effects of PATL2 depletion on the protein level of TUT7. In human PATL2-mutant oocytes or early embryos, the mRNA levels of TUT7 were comparable with control group (Fig. 7H). Similar with human PATL2 mutation, Patl2 knockdown using siRNAs did not induce the downregulation of Tut7 mRNAs in mouse oocytes (Fig. 7I). However, Patl2 knockdown induces the TUT7 protein decrease in mouse oocyte (Fig. 7J-K). Collectively, PATL2 is required for TUT7 protein level maintenance, and PATL2 probably collaborates with TUT7 to involve mRNA decay in human oocyte and early embryos.
PATL2 is required for global mRNA translation in oocytesWe observed that PATL2 overexpression resulted in higher protein levels of CDC23 and TUT7, while PATL2 knockdown led to a reduction in the protein levels of CDC23, APC1, MAD2L1, and TUT7 without affecting their mRNA levels. These findings suggest that, in addition to promoting protein stabilization through protein interactions, PATL2 may also maintain protein levels through regulation of mRNA translation. We examined the new protein synthesis ability by HPG (L-homopropargylglycine, an amino acid analog of methionine) incorporation. Consistent with the previous study [31], the HPG assay revealed that Patl2 knockdown resulted in a global decrease in mRNA translation in mouse GV oocytes (Figure S5A-B). In contrast, overexpression of human HA-PATL2 cRNAs in siPatl2 oocytes reversed PATL2 protein level and mRNA translation
留言 (0)