
記住我






Initially, we crossed heterozygous parents and collected mouse embryos ranging from embryonic day E11.5 to E14.5 (Table S1). After fixation, we recorded morphological features and performed genotyping. Subsequently, we sectioned the embedded embryos for hematoxylin and eosin (H&E) staining or immunofluorescence (IF) analysis. In our observation, the phenotypes of c-Jun+/+ and c-Jun+/− embryos were essentially identical. Among embryos at E11.5–E12.5, which exhibited normal size, no significant differences were observed in overall appearance or H&E staining (data not shown). However, embryos at E13.5–14.5 displayed distinct signs in c-Jun−/− embryos compared to their heterozygous or homozygous counterparts (c-Jun+/+, c-Jun+/−). These signs included diminished and discolored primary hepatic and heart regions (which was similar to previous reports) [18, 19], decreased transparency, increased whiteness of the brain region, and fading of surface blood vessels in some samples under transmitted bright light (Fig. S1A). Additionally, we occasionally observed smaller, white embryos, which appeared fragile, pale, and bloodless (Fig. S1A, H0150-8), consistent with previous reports of c-Jun KO embryos that ceased development and died [19, 20]. Genotyping confirmed these embryos as KO types, as expected, although their placentas appeared normal.
H&E staining analysis of sagittal sections of the CNS in E14.5 embryos revealed no significant differences in the cortex between WT and KO embryos (Fig. S1D-F). However, a noticeable decrease in cell density and sparse or loose nerve fibers was observed in the brain parenchyma of KO embryos (Fig. 1A, B, Fig. S1D-F). Particularly in the diencephalon region, WT embryos exhibited a normally developed thalamus with dense cells forming a heart-shaped outline (Fig. 1A, H015-1 c-Jun+/-) [21], Whereas, KO embryos from the same litter displayed a malformed thalamus (Fig. 1A, H015-3 c-Jun−/−). Similar malformations were observed in other litters (Fig. S1D-F). IF analysis further revealed that WT embryos had normal TUJ1+, CNTN2+ nerve fibers in the thalamus/diencephalon region, whereas these were significantly reduced in KO embryos (Fig. 1C). Staining for the thalamic characteristic marker TCF7L2 showed strong signals in WT embryos (Fig. 1D, a, c), but significant decreases (Fig. 1D, b) or undetectable levels (Fig. 1D, d) in KO embryos. IF staining revealed significantly reduced TUJ1 and TCF7L2 signals in KO embryos (Fig. 1E, F). To further verify nerve fiber loss, DiI (1, 1’-dioctadecyl-3, 3, 3’, 3’-tetramethylindocarbocyanine perchlorate) dye was used to examine dye diffusion along nerve fibers. In WT groups, DiI dye diffused smoothly along the mesencephalon and telencephalon (Fig. 1G). However, in KO groups, the dye gathered in the thalamic region with a truncated diffusion path (Fig. 1G), suggesting possible fiber loss and interruption of neural projections in the ventricular parenchyma. These findings suggest abnormal development of the thalamus/diencephalon in c-Jun KO mouse embryos, accompanied by nerve fiber loss in adjacent parenchyma.
Fig. 1
KO mouse embryos at E14.5 exhibited thalamic malformation in the diencephalon. (A, B) H&E staining of cJun+/- and KO embryos at E14.5 revealed sparse fibers in the ventricular parenchyma and malformation of the thalamus in the diencephalon of the KO embryo. The small inset in the upper-left corner of the image displays the global view of the embryo. H015-1: “H015” was the code number of the mother mouse, and “-1” was the embryo number in the litter, hereinafter the same. (C) IF staining of TUJ1 and CNTN2 showed that WT had normal nerve fibers in the thalamus/diencephalon region, while those of KO were broken down with scattered distribution. a–d, Scale bar: 100 μm. (D) IF staining of TCF7L2 showed strong signals in the thalamus/diencephalon region of WT (a, c), while it was significantly decreased (b) or undetectable (d) in the corresponding areas in the KO groups. Scale bar: 100 μm. (E, F) Statistical analysis of IF staining of TUJ1 and TCF7L2 in the thalamus/diencephalon region of mice embryo section at E14.5. The Y-axis represents the mean gray value of signals. n = 6, the values are presented as the mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001. (G) DiI dye diffusion in the adjacent areas of the diencephalon. The middle image shows a local magnification of the region near the thalamus. The arrows indicate the areas of apparent dye penetration obstruction in KO. The embryo H340-4 c-Jun-/- at the bottom-right corner was the “small and white” embryo, which was difficult to cut, thus the DiI crystals were implanted into the intact brain parenchyma area through an incision
Fig. 2
Loss of c-JUN in NPCs promoted neural stem cell induction and accelerated PAX6+cell differentiation. (A) Schematic of NPC induction. NIM: neural induction medium, NMM: neural maintain medium, SB: SB431542 (B) RT-qPCR gene expression test revealed that several NPC marker genes were significantly upregulated in the KO groups. (3 independent experiments). The values are presented as the mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001. (C) Flow cytometry plots illustrate the distribution of cells stained with PAX6 and NESTIN on day 12, 20, and 25. Differentiated PAX6−/NESTIN+ subcluster appeared in the KO groups, but were partially rescued in the OE groups on day 25. (D) Statistical analysis of the flow cytometry data showed a significant increase in cells within Q1 quadrant (PAX6−/NESTIN+ cells) (3 independent experiments). Two-way ANOVA. The values are presented as the mean ± SD, *p < 0.05, **p < 0.01, ***p < 0.001. (E) IF staining showed an increase in PAX6−/NESTIN+ rosettes (circled with yellow lines) in the KO groups on day 25. Scale bar: 50 μm. (F) Statistical analysis of IF staining showed a significant increase in PAX6−/NESTIN+ rosettes in the KO groups, which is partially reversed in the OE groups on D25 (n = 12, 4 independent experiments). *p < 0.05, **p < 0.01, ***p < 0.001
Fig. 3
Loss of c-JUN weakened the nerve fiber extension and adherent ability on plates in the neural culture. (A) Schematic of long-period continuous culture and thalamus neuron induction. NMM: neural maintain medium, TPM: thalamus patterning medium. (B) BF view of NPCs on day 28 (left panel) and day 43 (right panel). NPCs in the WT, KO, and OE groups could spread flat on the plates and extended plentiful fibrous axons on D28. In the KO groups, the cell bodies were more likely to aggregate and the axonal fibrous were significantly shrunk on day 43. Scale bar: 100 μm. (C) BF view of curled-up edges of cell colonies in the KO groups but not in the WT or OE group. Cells in the KO groups were easily detached from the culture dish (12-well dish) when subjected to enzymatic digestion. The white film indicated by the arrowheads shows cells shed after digestion. (D) Statistical analysis of the proportion of pro-detached colonies in NPCs on day 43. The KO groups showed a significantly high proportion of pro-detached colonies (3 independent experiments, 9 fields of view per group). *p < 0.05, **p < 0.01, ***p < 0.001. (E) IF staining of TUJ1 and TCF7L2 neurons on thalamus induction D1 and D7, respectively. In order to clearly mark the distribution of nerve fibers, TUJ1+ signals are highlighted by yellow lines (the rightmost column) Scale bar: 100 μm. (F) Statistical analysis of the ratio of the signal area of TUJ1 to TCF7L2 in NPCs on thalamus induction D7. The KO groups showed a significantly reduced ratio. This pattern was restored in the OE group. (3 independent experiments, 9 fields of view per group). *p < 0.05, **p < 0.01, ***p < 0.001, n.s.: not significant. BF, bright-light field
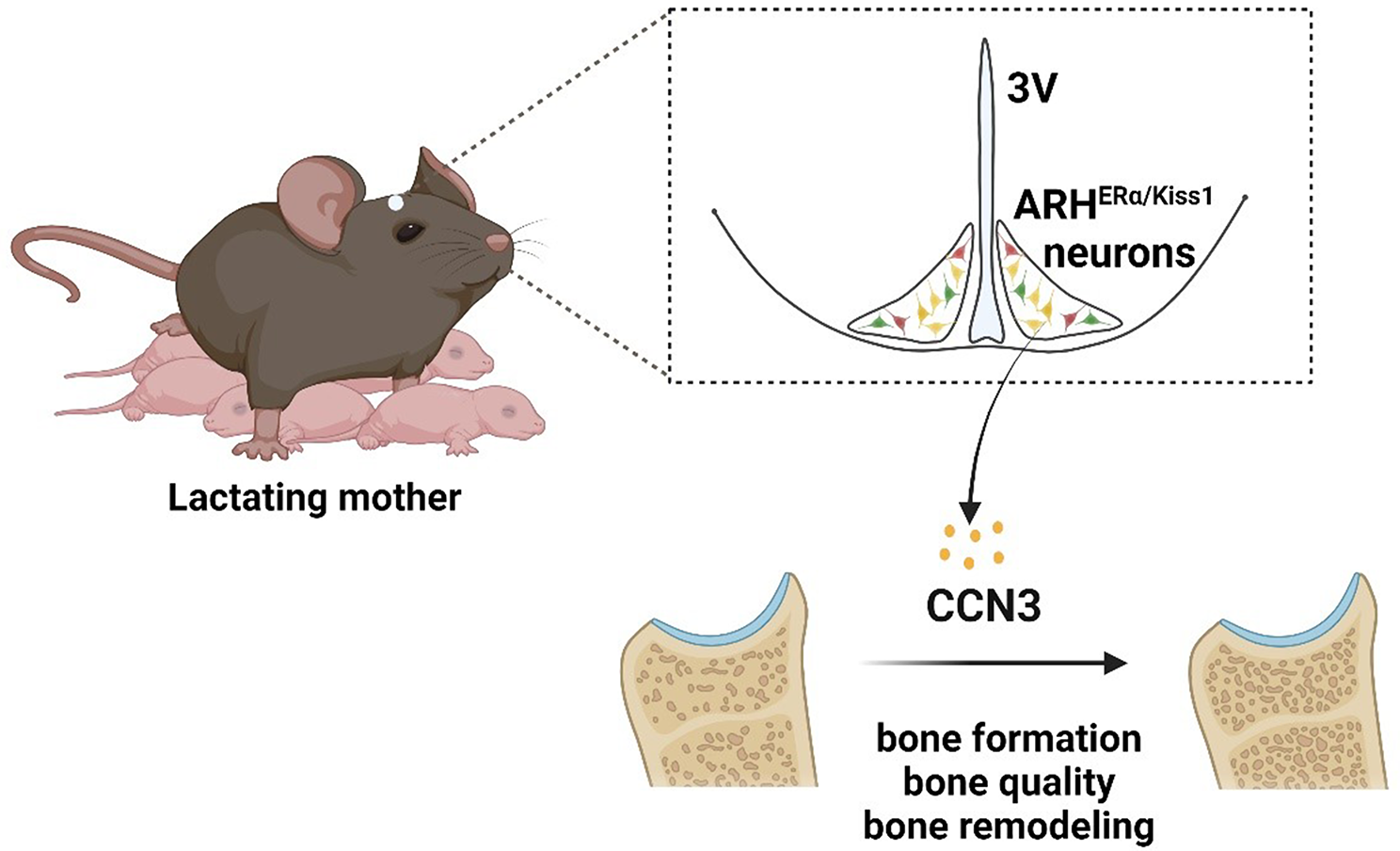
The temporal and spatial expression of c-Jun, Pax6 (a key gene in NPCs/neuron development), and Tcf7l2 (a thalamus-associated neural gene) was analyzed using the Mouse Organogenesis Spatiotemporal Transcriptomic Atlas (MOSTA) [21]. The data revealed the spatiotemporal expression patterns of these genes in mouse embryonic development from E9.5 to E16.5. IF staining on mouse brains at E14.5 confirmed their expression patterns as described by MOSTA (Fig. S2A, B). Our findings show that c-Jun is widely expressed in the embryonic development, with a stronger distribution in the subventricular zone in the forebrain cortex adjacent to diencephalon after E14.5. Pax6 expression is primarily detected in the ventricular lateral cortex of the forebrain and part of diencephalon, while Tcf7l2 is mainly found in the thalamic region of the diencephalon. Overlap in the expression of c-Jun and TCF7L2 was observed in the early developmental stage (E11.5 ~ 12.5) (Fig. S2C, D). The c-Jun KO may interfere with this relationship, contributing to the developmental abnormalities observed in the thalamus/dicephalus region.
Loss of c-JUN weakened the nerve fiber’s extensionTo investigate the role of c-JUN in human neural development, we generated c-JUN KO stem cell clones (KO#2, KO#10) from H1 ESCs using the CRISPR-Cas9 system, as previously described [12]. The karyotypes and cell proliferation/self-renewal of these KO clones appeared normal compared to WT H1 ESCs (Fig. S3A). Confirmation of c-JUN KO at the protein level was achieved through western blot analysis (Fig. S3B). Null expression of c-JUN mRNA in KO groups was verified by quantitative reverse transcription-polymerase chain reaction (RT-qPCR), whereas WT groups exhibited an upward trend of c-JUN expression throughout neural induction (Fig. S3C). During NPC induction (Fig. 2A), NESTIN and PAX6 signals were detected in both WT and KO H1 ESCs as early as day 4 with no significant differences. By around day 12, round, clustered neural rosettes began to appear, and by day 17, neural rosettes with a radial pattern expressing neural stem cell markers NESTIN and PAX6 were uniformly spread (Fig. S3E). After dispersing and replanting, typical cluster structures of rosettes were observed by day 23 (Fig. S3F). Notably, at the early stage (< day 20), complete depletion of c-JUN did not impede neural fate determination, and NPCs could generate typical PAX6+/NESTIN+ neural rosettes. Although rosette morphology showed no discernible differences before day 20, KO groups exhibited significantly increased expression of OTX1, PAX6, and FOXG1 tested by RT-qPCR (Fig. 2B). As the culture progressed, flow cytometry analysis at day 12, day 20, and day 25 identified a new sub-cluster appearing in the Q1 region in KO cells, indicating PAX6−/NESTIN+ cells (Fig. 2C), with the proportion of these cells significantly higher in KO than in WT on day 25 (Fig. 2D). This new sub-cluster was confirmed by IF staining, with some PAX6−/NESTIN+ rosettes observed in KO groups on day 25. These rosettes still exhibited NESTIN+ outlines, whereas internal PAX6 signals were significantly weakened or disappeared (Fig. 2E). Although PAX6−/NESTIN+ NPCs could occasionally be detected in the WT group, their proportion was much lower (Fig. 2F). Moreover, we established a tetracycline-inducible system to overexpress (OE) c-JUN using doxycycline (DOX). Western blot analysis confirmed the OE of c-JUN protein in KO cell lines (Fig. S3D). Upon OE of c-JUN in KO groups, the proportion of PAX6-/NESTIN + cells reduced to a level comparable to the WT group, and the percentage of PAX-/NESTIN + rosettes was also reduced to a level close to the WT group. These data indicate that the pro-differentiated NPCs were partially rescued (Fig. 2C-F). These findings suggest that c-JUN KO accelerates the induction of NPCs, and mature neural stem cells may be more prone to further differentiation.
When NPCs were cultured continuously (Fig. 3A), both WT and KO cells grew axons with typical neuronal morphology in the early days (D26–D35) (Fig. 3B, left panel). However, after day 35 without renewing the plate, WT cell axons continued to spread on the culture dish and extended plentiful fibrous axons that interwove with each other (Fig. 3B, right panel). In contrast, in KO groups, the axonal weaves significantly shrunk, and neuronal cell bodies tended to aggregate, with more cell colonies showing a tendency to curl up (Fig. 3B, right panel). We defined the neuronal colonies that were obviously aggregated with smooth edges and had a tendency to detach as “pro-detached colonies.” Statistical analysis of the proportion of pro-detached colonies in random field views revealed a significantly higher proportion in the KO groups (Fig. 3D). Moreover, when cells were subjected to enzymatic digestion (Accutase, STEM CELL, 07920) for 5 min at 37 °C, cells in KO groups detached more easily from the culture dish (Fig. 3C). This pro-aggregated and easier-detached phenomenon observed in long-period culture was rescued in OE groups (Fig. 3B-D). It is worth mentioning that neurons in all groups eventually aggregated and fell off, likely due to the exhaustion of coating proteins (e.g., laminin) in long-time cultures without changing plates. However, this process occurred significantly earlier in the KO groups. Furthermore, NPCs were induced into TCF7L2+ thalamus neurons using thalamus patterning medium (TPM) as used in ThO induction (Dulbecco’s Modified Eagle Medium [DMEM]/F12 medium containing N2, B27 without vitamin A, BMP7, PD325901, etc.) (Fig. 3A). Initially (day 21), newly seeded NPCs demonstrated undetectable TCF7L2 signals on cell bodies and weak class III beta-tubulin (TUJ1) signals along with short fibers (Fig. 3E, left panel). Following 7 days of induction, some cells expressed TCF7L2 signals along with abundant TUJ1+ nerve fibers (Fig. 3E, right panel). The ratio of TUJ1 signals to TCF7L2 signals was significantly lower in KO groups (Fig. 3F). This suggests that the TUJ1+ membrane and fibers (red signals) around the same number of TCF7L2+ neuron bodies (green signals) were significantly reduced in KO neurons. In 2D thalamus neuron differentiation, c-JUN KO restrained the initiation and extension of nerve fibers. Continuous culture of these TCF7L2+ thalamus neurons without renewing the plates in neural differentiation medium (the medium used in ThO induction in later stages) also revealed more aggregation and earlier shedding colonies in KO groups at day 43, similar to the phenomenon observed in long-period continuous NPCs culture. These findings collectively indicate that absence of c-JUN impairs nerve fiber extension and weakens neurons’ adhesive capabilities to the substrate, resulting in a propensity for neuronal colonies to aggregate and detach more readily from the culture plate.
COs derived from c-JUN KO hESCs exhibited robust exteriors but loose interiorsTo assess the impact of c-JUN on neural development within a model that closely mimics physiological conditions, we generated COs in an unguided manner (Fig. S4A). This whole-brain model exhibits preliminary ventral and dorsal axes and regional aggregation phenomena, with neural connections between the regions [22,23,24]. IF staining revealed the distribution of c-JUN in WT CO sections on day 12, 30 and 50 (Fig. S4B) and RT-qPCR test confirmed c-Jun expression increased during CO induction, while it was absent in the KO groups (Fig. S4C). In COs derived from H1ESCs, IF analysis detected neural stem cells markers (NESTIN, PAX6, SOX2, phospho-vimentin), neuronal marker (TUJ1), astrocytic marker (GFAP), and forebrain or cerebral cortex markers (FOXG1, TBR1, TBR2) within radially enlarged cavities and layered structures on day 30 and day 50. This suggests that the cultured COs exhibited cellular composition and morphogenetic patterns resembling early brain development (Fig. S4D-F) [24, 25]. Downregulation of stem cell-enriched genes (OCT4 and CK8) by day 16 and upregulation of nerve-related genes (PLAGL1 and TBR-2) from day 16 to day 30 were confirmed by RT-qPCR (Fig. S4G). Furthermore, at later stages, marker related to the forebrain cortex and synaptic function, such as TBR-1, CTIP-2, and SYNAP (D30–D60), were detected from day 30 to day 60 (Fig. S4H). These findings outlined a initial in vitro model of early cerebral development (Fig. S4I).
When comparing COs derived from WT and c-JUN KO cells, a more prominent transparent ring-shaped outer layer was observed in the KO on day 12–14 during CO culture (Fig. 4A-C). By day 24, an expanded transparent fan-shaped structure appeared in the KO, which was significantly wider than that in the WT group (Fig. 4D, E). IF analysis confirmed these observations during the corresponding period, showing that the transparent outer layer on day 12–14 was composed of PAX6+/NESTIN+ neural ectoderm (Fig. 4F). In COs derived from c-JUN KO cells, the neural ectoderm formed a robust closed ring on the outermost layer, whereas this ring was less robust and commonly interrupted in WT-derived COs (Fig. 4F). Subsequent structures derived from this ring in KO samples exhibited a significantly increased width of the PAX6+/NESTIN+ layer and enlarged cavities on day 30 (Fig. 4G). We further examined several neuronal and structural protein markers. Contactin protein 2 (CNTN2), which is predominantly expressed in the brain and encodes a glycosylphosphatidylinositol-anchored neuronal membrane protein involved in the proliferation, migration, and axon guidance of neurons during cerebral development [26, 27], was distributed in both the outer layer and some parts of the inner core in WT samples (Fig. 4H, a, b). However, in KO-derived COs, CNTN2 was primarily distributed in the outer cortical-like spheres (Fig. 4H, c-f). Similar distribution patterns were observed for the protein markers CLAUDIN5 and ZO1. In WT samples, CLAUDIN5 and ZO1 signals were detected in the outer layer as well as in the low-cell-density inner core (Fig. 4I, a, b, white arrows indicated). Conversely, in KO-derived COs, the signals were predominantly distributed in the cortex-like structures (Fig. 4I, c-f). Overall, RT-qPCR revealed the upregulation of neural marker genes such as LMO3, FOXG1, and TUJ1, whereas tight junction-related genes such as ZO-1 were downregulated in KO during CO induction (Fig. 4J).
Fig. 4
KO-derived CO showed a robust exterior, but less fiber-connected core. (A) Bright field view of CO induction from WT and KO cell lines., Scale bar: 200 μm for day 8–24; Scale bar: 500 μm for day 30–75. (B–C) KO groups showed a significant increase in the width of the neural ectoderm at day14 (n = 30, 3 independent experiments). *p < 0.05, **p < 0.01, ***p < 0.001. (D, E) The KO groups showed a significantly increased width of the expanded neural cortex-like structures at day30 (n = 30, 3 independent experiments). *p < 0.05, **p < 0.01, ***p < 0.001. (F, G) IF staining of CO on D14 and D30. The KO groups show robust PAX6+/NESTIN+ closed-ring shape ectoderm and increased width of PAX6+/NESTIN+ neural cortex-like layer (circled with yellow dashed line in the PAX6 channel). Scale bar: 100 μm. (H) IF staining of CNTN2 in COs at day 50. In WT, CNTN2 could be detected both in the periphery region and the interior (a, b). In the KO groups, CNTN2 was most distributed in the periphery but not in the core (c–f). Scale bar: 500 μm. (I) IF staining of ZO-1, CLAUDIN5 in COs at day 50. WT group showed tight-junction rich signals in the outer neural cortex-like layer as well as in the interior (indicated by the white arrows in b). In the KO groups, the signals were detected mostly on the outer layer, but not in the core. Scale bar: 500 μm. (J) qRT-PCR gene expression test revealed that several NPC/neuron marker genes (LMO3, FOXG1, and TUJ1) were upregulated while tight junction genes (ZO-1) were downregulated in the KO groups during CO induction (3 independent experiments). The values are presented as the mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001
These results indicate that COs derived from c-JUN KO cells exhibit a more robust neural ectoderm and extended subsequent neural structures but possess a less fibrous connected core. This trend aligns with the observations in c-JUN KO mouse embryos, where the cortex remained unchanged, whereas the ventricular parenchyma, particularly structures adjacent to the thalamus in the diencephalon, exhibited increased sparsity.
c-JUN KO upregulates nervous system development-related pathways and downregulates cell–cell adhesion pathwaysTo further understand the role of c-JUN in neural development, we conducted RNA-seq analysis on both WT and KO-derived COs at day 12 and day 24. Gene expression analysis using heat maps and principal component analysis (PCA) revealed significant differences between WT and KO samples at day 24 (Fig. 5A, B). Differentially expressed gene (DEG) analysis identified 141 genes, including MAP2, PAX6, DCX, and TBR1, that were significantly upregulated (p < 0.05, Fold change [FC] ≥ 2) in KO-derived COs compared to WT (Fig. 5C). Notably, other c-JUN family members JUNB, JUND, c-FOS, and FOSB exhibited no significant changes (data not shown). Gene Ontology (GO) analysis indicated that the upregulated genes in KO-derived COs were enriched in terms related to nervous system development, neuron migration, CNS development, and cerebral cortex development (Fig. 5E). This included genes involved in cerebral cortex development such as DCX, TBR1, and EMX2. Conversely, 223 genes, including RPS15A, CCNG1, FTL, TTR, and CDK1, were significantly downregulated (p < 0.05, FC ≥ 2) in KO-derived COs and were enriched in GO terms related to cell–cell adhesion, epithelial cell differentiation, and cholesterol homeostasis (Fig. 5C, D). The highest-ranking group, cell–cell adhesion, included genes such as TJP2 (also known as ZO-2), PERP, F11R, and DSG2, which play critical roles in maintaining tight cell connections. The volcano plot of DEGs in COs at D24 illustrated their distribution pattern and highlighted several genes (Fig. 5F). The up-regulated genes including NEDD4, NEDD4 E3 ubiquitin protein ligase, play critical roles in the regulation of a number of membrane receptors, endocytic machinery components and the tumor suppressor PTEN [28, 29]. TRH, thyrotropin releasing hormone, encodes a member of the thyrotropin-releasing hormone family. Thyrotropin-releasing hormone is involved in the regulation and release of thyroid-stimulating hormone, as well as prolactin. Deficiency of this hormone is associated with hypothalamic hypothyroidism [30, 31]. MAP2, microtubule associated protein 2, belongs to the microtubule-associated gene family and is thought to be involved in microtubule assembly, playing essential role in neurogenesis [32]. The down-regulated genes including POU5F1, POU class 5 homeobox 1, also known as OCT4, encodes a transcription factor containing a POU homeodomain that plays a key role in embryonic development and stem cell pluripotency. Aberrant expression of this gene in adult tissues is associated with tumorigenesis [33, 34]. CLDN6, claudin 6, a tight junctions protein, represents one mode of cell-to-cell adhesion in epithelial or endothelial cell sheets, forming continuous seals around cells and serving as a physical barrier to prevent solutes and water from passing freely through the paracellular space [35].
Fig. 5
c-JUN KO upregulated the pathways of nervous system development but downregulated the pathways in cell-cell adhesion. (A) Heat map of CO transcriptome sequencing on D12 and D24. (B) PCA analysis revealed the principal components clustered separately according to the two time points, and the divergence was obvious on D24 between the WT and KO groups. (C) Heat map of differentially expressed genes (DEG) in CO D24. (D, E) GO-BP analysis of DEG in CO D24. When compared to WT, the top 10 downregulated and top 10 upregulated biological pathways in KO are listed respectively. (F) Volcano map of DEG in CO day24
Previous studies have reported that c-JUN suppresses epithelialization [11, 36]. However, our GO analysis of COs at day 24 showed a group of epithelial marker genes were down-regulated (Fig. 5D). The neuroepithelium may be quite different from the general epithelialization or mesenchymal-to-epithelial transition. Factors such as cell types, differentiation stage, and cellular microenvironment can vary in gene functions and outcomes. We examined those 8 genes catalogued as “epithelial cell differentiation” in terms of significance ranking (p value), from high to low: F11R, LGALS3, KRT19, GSTA2, PGK1, CDK1, GSTA1, ELF3. Substantial evidence suggests they also play strong roles in structure maintenance and cells connection [37,38,39,40,41,42]. Thus, we hypothesize that in our neural developmental model, the down-regulated genes due to KO are likely associated with structural/cell adhesion pathways, and may differ from general epithelial differentiation and formation.
Overall, these data suggest that c-JUN functions as a regulatory hub closely associated with neurodevelopmental processes and the maintenance of cellular structure. c-JUN KO resulted in the upregulation of neurodevelopment-related pathways while downregulating cell–cell adhesion pathways in a model of cerebral development.
c-JUN KO affected the TCF7L2+ cell distribution in COs and mouse embryosTCF7L2 is known to be crucial for the development of forebrain and thalamus [43, 44], and it is commonly used as a marker specific for thalamus [45]. In embryos at E14.5 from the same litter H312, both WT embryos H312-7 (c-Jun+/+) and embryos H312-6 (c-Jun+/−) exhibited detectable TCF7L2+ signals in the ventral diencephalon (Fig. 6C, a, b, c). However, in the KO embryos, different staining patterns were observed: H213-5 (c-Jun−/−) displayed TCF7L2+ signals in the diencephalic area (Fig. 6C, d), whereas H312-9 (c-Jun−/−) and H312-10 (c-Jun−/−) showed hardly any detectable TCF7L2 signals in the expected areas (Fig. 6C, e, f). The TCF7L2 signals in another embryo at E14.5, H037-8 (c-Jun−/−), from a different litter, were also undetectable in the corresponding area (Fig. 6C, g). This discrepancy was partially mirrored in CO models: TCF7L2+ signals were present in WT-derived COs at day 50, particularly in subcortex-like regions and the outer margin of the core, reminiscent of the brain ventricular parenchyma, such as the diencephalon and thalamus (Fig. 6B, a, b). Conversely, TCF7L2 signals were scarcely detected in KO-derived COs (Fig. 6B, c, d). Analysis of 11 mouse embryos and 11 CO samples revealed that TCF7L2+ cells were detected in all WT samples, whereas clearly identifiable TCF7L2 signals were observed in only 50% of KO mouse embryos and 33% of KO CO samples (Fig. 6D, E). These findings indicate that abnormalities in the thalamus of the diencephalon may be traced back to aberrantly developed and distributed TCF7L2+ cells at the cellular level. The consistency between the trends observed in mouse embryos and COs suggests that CO models partially replicate differences observed in vitro.
Fig. 6
The disturbed expression pattern of thalamus/diencephalon marker TCF7L2 inc-Jun KO embryos and CO. (A) A diagram of the CO section and mouse embryo section; the red color area in the embryo section shows the thalamus in diencephalon. (B) WT-derived CO D50 could detect TCF7L2 signals, implying primary diencephanlic cells near the central area (a, b), while TCF7L2 was hardly detected in the KO groups. Image b is a partially enlarged image of the white arrow-indicated area in the image (a) Scale bar: 500 μm for a, c, d; 200 μm for (b) (C) WT embryos at E14.5 could detect TCF7L2 signals, implying thalamus in the diencephalon (a, b, c), while KO embryos appeared different
留言 (0)