
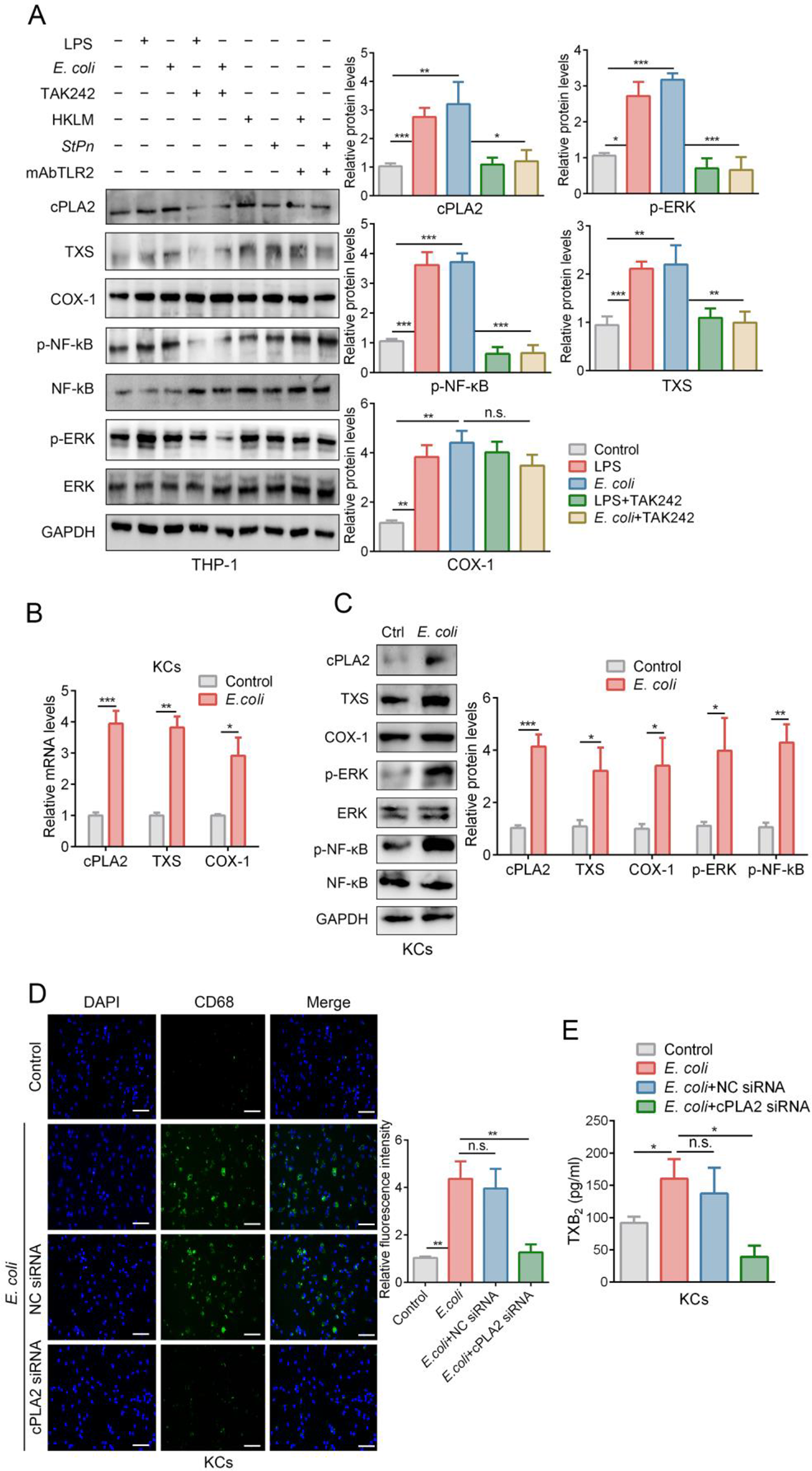
記住我







To investigate the effects of acetate in inflammation, we retrospectively examined the levels of acetate and inflammatory cytokines in serum of patients undergoing major abdominal gastrointestinal surgery (Clinical characteristics data see Supplemental Table 2). Our data demonstrated a linear relationship between serum acetate levels and IL-6 concentrations in these patients. In the Spearman correlation rank analysis, we found that acetate was positively correlated with IL-6 (r = 0.3733, p < 0.05) (Supplemental Fig. 1.1 A). As expected, comparing with the control group (infusion of 500 ml Ringer’s solution), serum acetate levels were significantly elevated 3 h after surgery in patients receiving acetate treatment (infusion of 500 ml Ringer’s acetate solution) (Fig. 1A). The abdominal surgery-induced upregulation of IL-6 was notably reduced in patients treated with acetate (Fig. 1B). Our findings also showed acetate significantly reduced the LPS-stimulated secretion of IL-6 and TNF-α in isolated CD14+ monocytes derived from healthy volunteers (Fig. 1C). We subsequently employed the CLP model, a systemic polybacterial infection model, to investigate the serum level of acetate. Our findings revealed a linear correlation between serum acetate levels and IL-6 in septic mice (r = 0.6351, p < 0.01) (Supplemental Fig. 1.1 B). Serum acetate levels were elevated 2.2-fold at 6 h and 4.3-fold at 24 h (Fig. 1D) after CLP. Interestingly, the serum acetate levels remained unchanged following the sole administration of exogenous acetate, while CLP treated with acetate significantly elevated the serum acetate levels (Supplemental Fig. 1.1 C, D). Moreover, acetate levels in major organs remain unchanged or exhibit a decrease (Supplemental Fig. 1E). These findings indicate that systemic inflammation plays a crucial role in the induction of intrinsic acetate.
Fig. 1
Acetate is Increased in serum during inflammation and exogenous acetate can suppress immune response and protect mice from CLP sepsis. A Acetate levels in serum of patients undergoing major abdominal surgery involving gastrointestinal tract received ringer’s acetate (20 ml/kg) infusion as compared to normal ringer’s (Ctrl) 3 h after the injection. B IL-6 level in serum of patients undergoing major abdominal surgery vein injected with ringer’s acetate as compared to normal ringer’s (Ctrl) 3 h after the injection was analyzed by ELISA. C Gene expression of TNF-α and IL-6 of CD14+ monocytes isolated from healthy human PBMCs stimulated with 6 h LPS (100ng/ml) in the presence of acetate (10 mm) were analyzed by qRT-PCR. D Acetate levels in serum of C57BL/6 mice 6 h and 24 h after CLP. E Mice were intra-peritoneal injected with acetate immediately after the establishment of CLP model at the indicated dose, and the mortality of CLP mice was detected (n = 20). Kaplan–Meier curves and the log-rank test were used to compare mortality rates, *p < 0.05. F Pro-inflammatory cytokines in serum of mice treated with acetate as in (E) 6 h after CLP was analyzed by ELISA. G Lung (Scale bars, indicated) sections from mice at 24 h after sham or CLP surgery treated with acetate at indicated dose were analyzed by Hematoxylin and Eosin staining (H&E staining). H Mice received an intra-peritoneal injection of acetate (500 mg/kg) immediately, 2 h, 4–6 h after the establishment of CLP model, and the mortality of CLP mice was detected (n = 20). Kaplan–Meier curves and the log-rank test were used to compare mortality rates, *p < 0.05 I BMDMs were treated with indicated dose of acetate prior to LPS exposure. 6 h later, TNF-α and IL-6 were analyzed by ELISA. J Gene expression of TNF-α and IL-6 of acetate pretreated BMDMs in indicated time exposure to LPS were analyzed by qRT-PCR. K BMDMs were treated with CpG (1 µM) or transfected with poly(I: C) (1 µg/ml) for 24 h in the presence of 10 mm acetate. TNF-α and IL-6 in corresponding cell culture supernatants were analyzed by ELISA. L Representative Western blot of MAPKs pathway-related proteins, phosphor- and total p65 levels in LPS stimulated BMDMs pretreated with acetate. Statistical analysis was performed by Student’s t-test and one-way ANOVA. The survival rates were analyzed by the LogRank test. Data are expressed as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001 as indicated. Data are pooled from at least two independent experiments
In addition, we observed that the exogenous administration of acetate significantly improved the survival rates of CLP mice in a dose-dependent manner (Fig. 1E)., reduced the expression of inflammatory factors (TNF-α, IL-6 and IL-1β) in serum after 24 h of CLP (Fig. 1F), decreased hepatic and renal functions markers (ALT and BUN) (Supplemental Fig. 1.1 F, G), and improved the pathology of lung (Fig. 1G). The analysis of cell counting changes in the inflammatory infiltrating cells within the livers of CLP mice (Supplemental Fig. 1.1 I, J) revealed no significant differences between the acetate-treated group and the sham group. Administration of acetate at a concentration of 50 mM in drinking water for three days prior to CLP significantly enhanced survival rates (Supplemental Fig. 1.1 K). More importantly, the intraperitoneal administration of acetate at 2/4 h post-CLP still significantly improved the survival rates (Fig. 1H). The data indicate the potential preventive and therapeutic effects of acetate in the context of sepsis. In BMDMs. We observed that acetate concentrations as low as 10 mM significantly inhibited the production and expression of pro-inflammatory cytokines (Fig. 1I, J). Notably, no significant changes in cell viability were detected even at a concentration of 100 mM (Supplemental Fig. 1.2 A). Furthermore, both acetate and acetic acid demonstrated a dose-dependent inhibition of inflammatory cytokine production induced by LPS (Supplemental Fig. 1.2 B, C). The pH of the culture media (data not shown) remained unchanged with the lowest dose of acetate (10 mM), which was used for subsequent research.
Sepsis can occur as a result of various bacterial, viral, or fungal infections [24]. To further explore the effects of acetate on other TLR signaling pathways, we stimulated BMDMs with poly(I:C) (a ligand for TLR3) and CpG (1 µM; a ligand for TLR9). Acetate significantly inhibited cytokine production induced by all three ligands (Fig. 1K), indicating that its effects are not limited to TLR4. These findings suggest that acetate may have a broader role in the context of both bacterial and viral infections.
Ligation of TLR4 results in the activation of nuclear factor κB (NF-κB) and mitogen-activated protein (MAP) kinases, ultimately promoting cytokine transcription. The inhibitory effect of acetate on NF-κB phosphorylation was observed as early as 15 min (Fig. 1L), while the MAP kinase pathways (ERK, JNK, and p38) in BMDMs remain unchanged (Supplemental Fig. 1.2 D). Furthermore, we observed that the LPS induction of M2 polarization genes, Arg1, Rentnla and Mgl2 (Supplemental Fig. 1.2 E), in acetate-pretreated BMDMs were unaffected compared to LPS group, which implied that acetate exposure may not be able to switch the polarization of macrophage. These data suggest that acetate ameliorates inflammatory cytokine production and negatively regulates NF-kB signaling in macrophages.
Acetate ameliorate inflammatory response through glycolysis attenuationMetabolic rewiring from oxidative phosphorylation toward glycolysis is essential for mounting efficient pro-inflammatory responses in macrophages. It has been suggested that acetate can regulate glucose metabolism [5, 24]. To further investigate the role of glucose metabolism in the immuno-suppressive effect of acetate in macrophages, we first assessed glucose uptake in the culture medium of BMDMs. The glucose levels in the supernatant increased upon stimulation with lipopolysaccharide (LPS), whereas treatment with acetate resulted in a slight yet significant reduction in extracellular glucose levels (Fig. 2A). Accordingly, lactate production in culture medium decreased in acetate-treated macrophages upon LPS stimulation (Fig. 2A). Next, we used the fluorescent glucose analog 2-NBDG to measure glucose uptake in BMDMs. In line with the increased lactate secretion, glucose uptake was significantly decreased in acetate-treated BMDMs after 0.5 h treatment with LPS (Supplemental Fig. 2A). To investigate the glucose uptake in septic mice, 2-NBDG was intravenously injected after challenge of LPS. Injection of LPS resulted in a mild increase in the uptake of 2-NBDG by splenic macrophages. However, acetate-treated mice showed a lower uptake of 2-NBDG (Fig. 2B). These findings indicate that acetate dampened LPS-driven glucose uptake both in vitro and in vivo. Additionally, we found a decrease in essential glycolysis enzymes such as PFKFB3, PKM2 and the main glucose transporter, Glut1 (Fig. 2C, Supplemental Fig. 2B). These results indicate that acetate suppressed the metabolic shift toward glycolysis in LPS-stimulated macrophages.
Fig. 2
Glycolysis is essential in anti-inflammatory effects of acetate in macrophages. A The lactate and glucose levels of the culture supernatants of acetate pretreated BMDMs stimulated with 100 ng/ml LPS for 6 h were assayed by colorimetric assay. Uptake and export were calculated from the difference between controls and activated samples. B Mice received 2-DG (2 g/kg) or vehicle control intravenously 30 min before receiving LPS (5 µg) intravenously; acetate (10 mM) was i.p administrated for 0.5 h before LPS stimulation;1 h later, 2-NBDG (50 µg) was injected and 15 min later mice were killed and splenic Gr1−CD11b+F4/80+ macrophages were analyzed by flow cytometry for the uptake of 2-NBDG. C Representative Western blot analysis of glycolysis pathway-related proteins, Glut1, PKM2 and PFKFB3 in acetate pretreated BMDMs under LPS exposure for indicated time. D, E The ECAR of acetate pretreated BMDMs stimulated with or without 100 ng/ml LPS for 1 h was assessed by the Seahorse assay before and after sequential addition of glucose, oligomycin and 2DG. F, G The OCR was assessed by the Seahorse assay before and after sequential addition of oligomycin, FCCP, rotenone, and antimycin. H, I Acetate pretreated BMDMs were stimulated by 100ng/ml LPS for 6 h in the presence of IFN-γ (100ng/ml) or GM-CSF (100ng/ml). TNF-α and IL-6 in corresponding cell culture supernatants were analyzed by ELISA. Statistical analysis was performed by one-way ANOVA. Data are expressed as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001 as indicated. Data are pooled from at least two independent experiments
We further assessed the effect of acetate on the glucose metabolism of macrophages. The glycolytic and mitochondrial metabolic activity was measured using a Seahorse analyzer. Results from the oxygen consumption rate (OCR) and extracellular acidification rate (ECAR) demonstrated that upon 1 h stimulation of LPS, supplementation of acetate resulted in a significant decrease in glycolysis (Fig. 2D, E), whereas OCRs were
unchanged compared to medium control (Fig. 2F, G). Acetate (10 mM) alone greatly enhanced mitochondrial OXPHOS, as evidenced by increased basal respiration as early as 1 h after treatment (Fig. 2G). Overall, our results indicate that glycolysis was suppressed in acetate-treated macrophages upon immune stimulation.
Next, to investigate whether the glycolysis suppression was essential for the effect of acetate, we used IFN-γ or GM-CSF, which have been shown to enhance glycolytic activity in vitro and in vivo [25, 26]. Pre-incubation with IFN-γ or GM-CSF (Fig. 2H, I) resulted in nearly completely abrogating the effect of acetate on LPS-induced production of cytokines. As expected, decreased LPS-driven mRNA expression of Glut1, PFKFB3, HK2, and MCT4, as well as the flow cytometry analysis of Glut1 of BMDMs by acetate, were all reversed by IFN-γ or by GM-CSF (Supplemental Fig. 2C-F), indicating the enhancement of glycolysis could reverse the anti-inflammatory effect of acetate. Together, these findings suggest that acetate acts as a key modulator of the immune response of macrophages by affecting glycolysis.
Stabilization of Hif-1α reverse the anti-inflammatory effect of acetate in vitro and in vivoTo elucidate how acetate mediates decreased glycolysis in BMDMs, we measured the expression levels of the AKT/mTOR/ Hif-1α pathway [27] which regulates glucose metabolism. Acetate dampened the expression of HIF-1alpha after LPS stimulation in BMDMs (Fig. 3A) and the overexpression of Hif-1α abrogated the suppression of IL-6 and TNF-alpha induced by acetate in RAW264.7 cells (Fig. 3B). Our results showed that the phosphor- and total AKT remains unchanged in the absence or presence of acetate. The phosphoredmTOR was decreased in acetate treated macrophage 15 min after LPS stimulation. (Supplemental Fig. 3A, B). In the presence of mTOR pathway activator MHY1485 [28] and 3BDO which enhanced the expression of p70S6K, the downstream target of mTOR (Supplemental Fig. 3C), the suppression of IL-6 and TNF-alpha induced by acetate remains unaltered (Supplemental Fig. 3D). Meanwhile, mTOR inhibitor rapamycin did not reverse the immuno-suppressive effect of acetate (Supplemental Fig. 3D), which suggesting a less possibility of involvement of mTOR axis in the immuno-suppressive effect of acetate. Thus, these data suggeste that Hif-1α, but not AKT/mTOR, regulate the immuno-suppressive effect by acetate.
Fig. 3
Stabilization of HIF1α reverse the anti-inflammatory effect of acetate by increasing glycolysis in vitro and in vivo. A Representative Western blot of HIF1α in acetate pretreated BMDMs under LPS exposure for indicated time. B RAW264.7 cells were transduced with vector specific for Hif-1α and control vector for 24 h and treated with 10mM acetate for 0.5 h, then stimulated with LPS for 6 h, TNF-α and IL-6 in supernatant were determined by ELISA. C Representative Western blot of Hif-1α in acetate pretreated BMDMs under LPS exposure for indicated time in the presence of MG132 (10 µM). D Representative Western blot of Hydroxyl-Hif-1α in acetate pretreated BMDMs under LPS exposure for indicated time. E BMDMs were stimulated with acetate prior to LPS for 6 h in the presence with Hif-1α stabilizer DMOG (0.5mM). TNF-α and IL-6 in corresponding cell culture supernatants were analyzed by ELISA. F Representative Western blot of pp65/p65 in acetate pretreated BMDMs under LPS exposure in the presence of DMOG (0.5mM) for indicated time. G Mice treated with or without DMOG (320 mg/kg, i.p) 3 h before CLP. Acetate (500 mg/kg i.p) was given immediately after CLP. 12 h after CLP, TNF-α, IL-6 and IL-1β in serum were analyzed. H AST and BUN in plasma 12 h after CLP mice treated as in (F). I Survival of CLP mice treated as (G) (n = 8/group. p = 0.0390 for CLP + PBS versus CLP + A; p = 0.8319 for DMOG + CLP versus DMOG + CLP + A). Statistical analysis was performed by one-way ANOVA. The survival rates were analyzed by the LogRank test. Data are expressed as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001 as indicated. Data are pooled from at least two independent experiments
Degradation of HIF-1α by the proteasomes is tightly regulated by its prolyl hydroxylation [28]. Since the mRNA level of Hif-1α was not decreased in acetate treated BMDMs (Supplemental Fig. 3E), we asked whether acetate influences the degradation of Hif-1α protein which is proteasome dependent. We found that proteasome inhibitor MG132 abolished acetate-mediated Hif-1α decrease, as well as the cytokine suppression (Fig. 3C, Supplemental Fig. 3F). Furthermore, the expression of hydroxyl-Hif-1α was increased as early as 15 min in acetate treated cells (Fig. 3D). Also, we found that DMOG [29], a competitive inhibitor of Hif-1α prolyl hydroxylase that stabilize the Hif-1α, abolished acetate-mediated alteration in cytokine production (Fig. 3E), phosphor- and total p65 (Fig. 3F), glucose uptake and lactate excretion (Supplemental Fig. 3G) as well as the mRNA level of glycolysis-related enzymes (Supplemental Fig. 3H). We then injected mice with DMOG before CLP and acetate treatment to evaluate the essential role of hydroxyl-Hif-1α in vivo. Our results showed that the DMOG abrogated the acetate-mediated alleviation of the pathology of liver and lung (Supplemental Fig. 3I). Similarly, DMOG reversed the decreased TNF-α, IL-6 and IL-1β in serum (Fig. 3G) and restored the decreased hepatic and renal functions marker AST and BUN (Fig. 3H) induced by acetate compared with control mice. Moreover, DMOG significantly impaired the prolonged survival in acetate treatment CLP mice (Fig. 3I). Thus, our result suggest acetate may ameliorate inflammatory cytokine production through increasing the proline hydroxylation and subsequent degradation of Hif-1α.
Acetate promotes histone acetylation but decreased acetyl-transfer to p65Exogenous acetate can be transported through transmembrane diffusion or signal via GPR41 or GPR43 [30]. We and other researchers have found GPR43 has a crucial role in the protection of acetate on innate immune responses, including those of macrophages and neutrophils. Unexpectedly, mRNA of GPR41 and GPR43 was found downregulated in acetate-exposed BMDMs (Supplemental Fig. 4A), and neither the knockdown of GPR43 or GPR41, separately or in combination, reversed the effect of acetate on LPS-induced production of cytokines (Supplemental Fig. 4B, C), suggesting that GPR43 and GPR41 may not be involved in the anti-inflammatory effect of acetate. Next, we found an increased uptake of acetate by macrophages exposed to LPS and exogenous acetate for 6 h (Fig. 4A). After cellular uptake, acetate is quickly converted into acetyl-CoA, as our result showed that cellular acetyl-CoA transiently increased in 30 min after LPS exposure in BMDMs pretreated with acetate (Fig. 4B). There are two enzymes that have proved to be capable of using acetate as a substrate: acetylCoA synthetase 1 (ACSS1) and ACSS2 [31]. Apart from ACSS1 and ACSS2, ATP Citrate Lyase (ACLY) also accounts for the acetyl-CoA pool expanding in acetate treated memory CD8+ T Cells [32]. Thus, we measured ACSS1, ACSS2 and ACLY expression and found LPS induced significant increase of ACSS1, ACSS2 and ACLY, however only ACSS2 increased in acetate combined LPS stimulation (Fig. 4C). Furthermore, an ACSS2 inhibitor (Fig. 4D) or knockdown of ACSS2 (Fig. 4E), but not knockdown of ACSS1 or ACLY (Supplemental Fig. 4D, E), reversed the effect of acetate on LPS-induced production of cytokines, supporting the notion that ACSS2 involved in the anti-inflammatory effect of acetate. Together, these results suggeste transmembrane-diffused acetate synthesized to acetyl-CoA by ACSS2 participates in anti-inflammatory effects.
Fig. 4
Protein acetylation is involved in immuno-suppressive effect of acetate. A BMDMs were treated with acetate prior to LPS (100 ng/ml) exposure for 6 h, acetate levels were measured in the cell supernatant by colorimetric assay. Changes were calculated from the difference between controls and activated samples. B Acetyl-CoA concentration in in BMDMs treated with LPS for indicated time in the presence of acetate determined using a fluorimetric assay. C Western blot analysis of acetate-acetyl-CoA pathway-related proteins, ACSS1, ACSS2 and p-ACLY /ACLY in acetate pretreated BMDMs under LPS exposure for indicated time. D BMDMs were stimulated with acetate prior to LPS for 6 h in the presence with indicated dose of ACSS2 inhibitor. TNF-α and IL-6 in corresponding cell culture supernatants were analyzed by ELISA. E PMs were treated with si-ACSS2 for 48 h following combined stimulation, and the indicated cytokine levels were analyzed by ELISA. F Acetate increased H3K9, H3K18 and H3K27 acetylation levels. BMDMs were treated with or without acetate (10 mM) in the presence of LPS for indicated time. Hstones were extracted. The Global histone acetylation levels were determined by western blot. Total H3 served as a loading control. G Representative Western blot analysis of H4K5, H4K8 and H4K12 acetylation levels in acetate (10 mM) pretreated BMDMs under LPS exposure for indicated time. Total H4 served as a loading control. H Representative Western blot analysis of Histone Deacetylase SIRT1/3/6 and HDAC1/3/4 in acetate (10 mM) pretreated BMDMs under LPS exposure for indicated time. I BMDMs were stimulated with acetate prior to LPS for 6 h in the presence of Gacinol (10 µM). TNF-α and IL-6 in corresponding cell culture supernatants were analyzed by ELISA. J Representative Western blot analysis of pp65 and HIF-1α levels in acetate (10 mM) pretreated BMDMs under LPS exposure for indicated time in the presence of Gacinol (10 µM). K BMDMs were stimulated with acetate prior to LPS for 6 h in the presence of TSA(50 nM). TNF-α and IL-6 in corresponding cell culture supernatants were analyzed by ELISA. L Representative Western blot analysis of pp65 and HIF-1α levels in acetate (10 mM) pretreated BMDMs under LPS exposure for indicated time in the presence of TSA (50 nM). M Representative Western blot analysis of Ac-NF-κB p65 Lys310 in acetate (10 mM) pretreated BMDMs under LPS exposure for indicated time. N 200 µg of protein lysates of acetate (10 mM) pretreated BMDMs under 1 h LPS exposure were precipitated with anti-p65 antibody and probed for acetylation. showing acetate-induced acetylation in p65 immunocomplex. Statistical analysis was performed by Student’s t-test and one-way ANOVA. Data are expressed as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001 as indicated. Data are pooled from at least two independent experiments
We next investigated the subsequent effects of the increased acetyl-CoA. As ACSS2-produced acetyl-CoA is the substrate for protein acetylation, we assessed the impact of acetate on acetylation. Increased acetyl-CoA usually promotes the histone acetylation in nucleus [33]. We found acetate rapidly promoted the level of H3K9ac, H3K18ac as well as H3K27 in LPS-stimulated and unstimulated macrophages within 15 min (Fig. 4F), (Supplemental Fig. 4F). Consistently, histone 4 (H4) acetylation at lysine K8, K5 and K12 was markedly enhanced by acetate exposure relative to control macrophages. (Fig. 4G), (Supplemental Fig. 4G). Although global histone acetylation was promoted, we found acetyl transferase (CBP) and lysine de-acetyl transferase (HDAC1,3,4 and Sirt1,3,6) were all downregulated in the acetate treated BMDMs compared with LPS-stimulated macrophages (Fig. 4H) (Supplemental Fig. 4H, I). Moreover, neither garcinol, an inhibitor of acetyltransferases [34], nor TSA, an inhibitor of de-acetyl transferase, could reverse acetate-mediated cytokine suppression or the decreased phosphor-p65 and Hif-1α (Fig. 4I-L). And the knockdown of Sirt1/3/5 (Supplemental Fig. 4J) also showed limited influence on acetate effects. These results suggested histone acetylation, though enhanced by acetate treatment, is not essential for acetate-mediated suppression of cytokine production.
Histone acetylation forms an acetyl-pool for acetyl-cycle for histone and non-histone modifications. As acetate decreased acetyl transferase, this may suppress the non-histone acetylation. Indeed, we found acetate treatment decreased the acetylation of Lys310 (Fig. 4M), a most-studied acetylation residue of p65 [35, 36]. And the total acetylation of p65 was also decreased (Fig. 4N). Previous study reported the enhanced acetylation of p65 promoted the inflammatory response in macrophage. Thus, these results suggested acetyl-modification of p65 may involve in acetate-mediated anti-inflammation.
Together, these results show that the acetate enhanced the histone acetylation but suppressed the acetyl-transfer to p65.
Lipid metabolism may not involved in the immuno-suppressive effect of acetateActivation of the de novo lipogenesis and cholesterol biosynthesis pathways is also crucial for the inflammatory response to LPS [37,38,39,40]. After ligation with CoA by ACSS2, acetate can be converted to acetylCoA, which could be carboxylated by acetyl-CoA carboxylase1(ACC1) and condensated by fatty acid synthase (FASN) into fatty acids (Fig. 5A). To evaluate the role of lipid metabolism, we measured the key genes involved in de novo lipogenesis (ACC and FASN) and found ACC and FASN were induced in mRNA level in acetate-treated BMDMs (Fig. 5B) (Supplemental Fig. 5A). Next, we assessed the free fatty acid level by LC-MS. While LPS treatment significantly increased free fatty acid levels in control macrophages, we observed no changes in free fatty acid levels in acetate pretreated BMDMs compared to LPS group (Supplemental Fig. 5C). Moreover, the knockdown of ACC or FASN (Fig. 5C), or the administration of C75, an inhibitor of FASN (Fig. 5D), failed to reverse the cytokine suppression induced by acetate. Thus, these data suggested the immuno-regulatory effect of de novo lipogenesis was not found observed in this setting.
Fig. 5
Fatty acid synthesis is not involved in the immuno-suppressive effect of acetate. A Schematic showing fates of acetyl-CoA in the de novo lipogenesis and sterol biosynthesis pathways. B Relative mRNA expression of ACC and FASN in acetate pretreated BMDMs exposure to LPS for 1 h were analyzed by qRT-PCR. C PMs were treated with si-ACC or si-FASN separately for 48 h following combined stimulation, and the indicated cytokine levels were analyzed by ELISA. D BMDMs were stimulated with acetate prior to LPS for 6 h in the presence of FASN inhibitor C75 (10 µM). TNF-α and IL-6 in corresponding cell culture supernatants were analyzed by ELISA. E BMDMs were stimulated with acetate prior to LPS for 6 h in the presence of HMGCR inhibitor Fluvastatin (10 µM). TNF-α and IL-6 in corresponding cell culture supernatants were analyzed by ELISA. F PMs were treated with si-CPT1a for 48 h following combined stimulation, and the indicated cytokine levels were analyzed by ELISA. Statistical analysis was performed by Student’s t-test and one-way ANOVA. Data are expressed as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001 as indicated. Data are pooled from at least two independent experiments
In the mevalonate pathway of cholesterol synthesis, Hydroxy-3-methylglutaryl-CoA synthase 1 (HMGCS1) is the key enzyme. We found fluvastatin, the inhibitor of HMGCS, cannot reverse the suppressive effect of acetate (Fig. 5E). Furthermore, while the expression of key enzyme of fatty acid oxidation, carnitine palmitoyl transferase 1 A (CPT1A) was upregulated (Supplemental Fig. 5B), the knockdown of CPT1A failed to reverse the suppressive effect of acetate (Fig. 5F). Together, our current data suggest that the lipid metabolism in macrophages stimulated by LPS for 6 h was not changed by acetate, and lipid metabolism might not be involve in the immuno-suppressive effect of acetate.
留言 (0)