
記住我



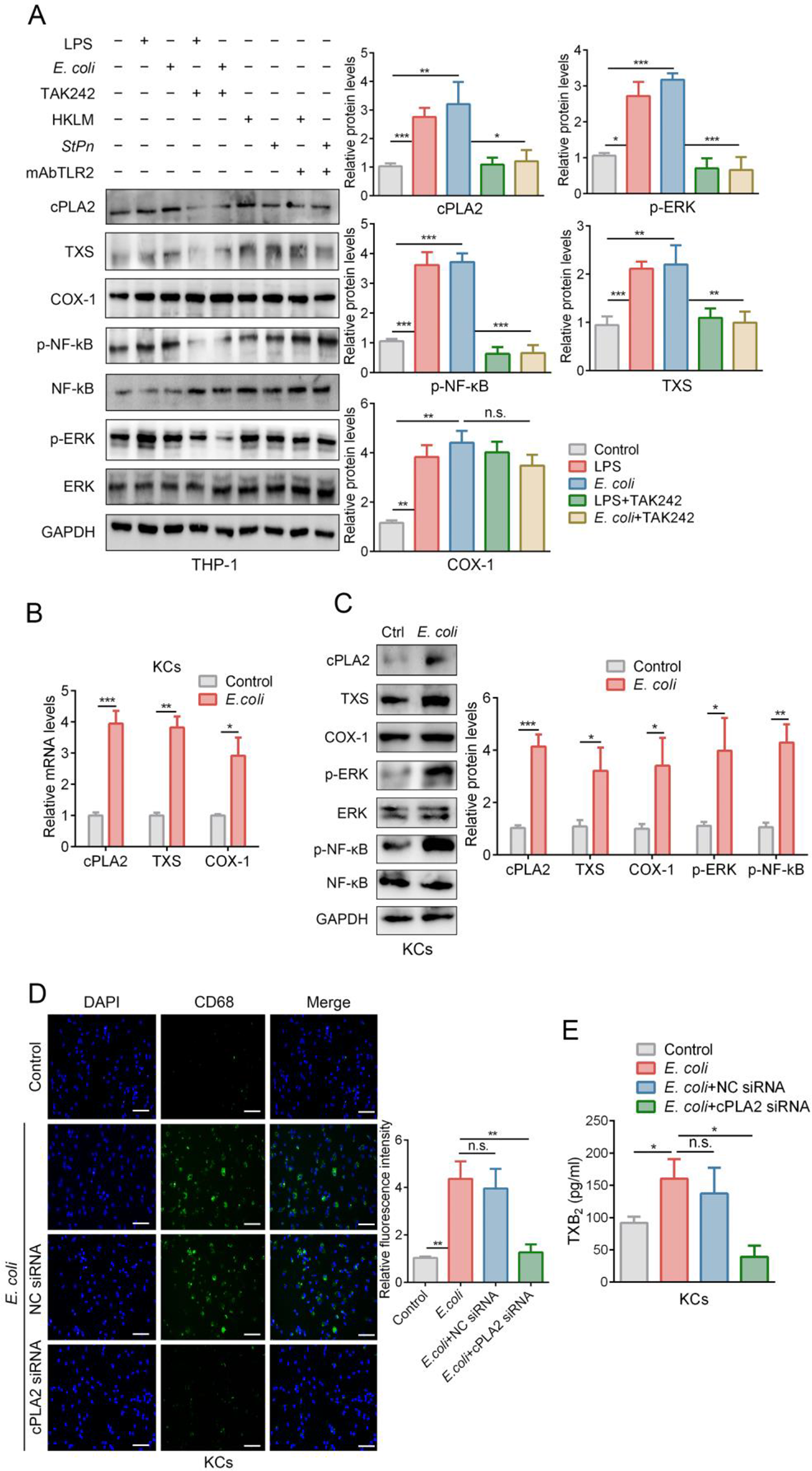




We compared the effects of increasing doses of amiloride for 72 h in hormone-sensitive LNCaP cells and their castration-resistant derivative C4-2 cells, as well as in an unrelated hormone-insensitive cell line CWR22Rv1 (denoted henceforth as 22Rv1). We have previously shown that all three cell lines express abundant ErbB3, HER2 and EGFR protein and mRNA (but not ErbB4) [23, 24]. Amiloride has been used at concentrations up to 1 mM in numerous cancer cell lines, including PCa [30, 46], but we tested the response of ErbB family kinases at amiloride concentrations only up to 100 µM. In LNCaP cells, baseline EGFR and HER2 were mostly cytoplasmic, with EGFR levels decreasing steadily until 75 µM, and then abruptly increasing at 100 µM (Fig. 1A). ErbB3, in contrast, displayed both cytoplasmic and nuclear localization at baseline; however, with increasing amiloride the ratio of cytoplasmic to nuclear ErbB3 steadily increased, until at 75µM amiloride it was significantly cytoplasmic. This result was validated using immunofluorescence microscopy which showed nuclear ErbB3 in vehicle-treated cells but not in 75 µM amiloride-treated cells (Fig. 1B). Surprisingly, at 100 µM amiloride, the levels of nuclear ErbB3 appeared to be restored, and the levels of cytoplasmic EGFR appeared to be increased. Lower doses of amiloride were examined as well and demonstrated smaller changes in localization in LNCaP cells (Fig. 1C); however, it is obvious that at 25 µM, ErbB3 translocated from a nuclear location to a cytoplasmic location, while at 10 µM, the level of nuclear ErbB3 is somewhat reduced. In parallel, amiloride also caused a dose dependent inhibition in cell growth, as indicated by MTT assay, with an IC50 = 18.07 µM (Fig. 1D). Significant suppression of cell growth was observed at 25 µM (p = 0.0236), and higher doses, suggesting correlation between loss of cell viability vs. loss of nuclear ErbB3 localization.
Fig. 1
Amiloride promoted ErbB3 translocation from the nucleus to the cytoplasm and the plasma membrane in HSPC cells (A) Hormone-sensitive LNCaP cells were treated with varying concentrations of amiloride for 72 h before being lysed, fractionated and analyzed by immunoblot. (B) Immunofluorescence microscopy (IF) in LNCaP cells treated with DMSO or 75 µM or and probed with IF specific C-terminal ErbB3 antibodies or (C) 10µM or 25µM amiloride and probed with IF specific N-terminal ErbB3 antibodies for 72 h (scale bars = 30 μm). Note that vehicle treated LNCaP cells expressed nuclear ErbB3 (red) whereas amiloride-treated cells had significantly decreased ErbB3 expression in the nucleus (hollowed out). Location of nuclei are identified by blue DAPI staining. Plasma membrane localization of ErbB3 at cell-cell junction was also noted in amiloride-treated but not in vehicle treated cells. Note that both N- and C-terminal ErbB3 antibodies demonstrate lighter nuclear staining in amiloride-treated cells. (D) Cells were subjected to viability assays using the stated concentrations of amiloride. p-values are calculated with respect to DMSO
In contrast, in C4-2 cells, all ErbB receptor tyrosine kinases, including ErbB3, were overwhelmingly cytoplasmic, even at baseline (Fig. 2A). Cell viability still showed a dose-dependent decrease with increasing amiloride concentration causing half-maximal inhibition at 22.41 µM, indicating a different mode of action of amiloride in these cells (Fig. 2B). When the CRPC cell line 22Rv1 was similarly examined, there was a dose-dependent decrease in EGFR, but a dose-dependent increase in HER2 with increasing amiloride, while ErbB3 levels were not significantly affected (Fig. 2C). As a result, 22Rv1 cells were significantly less susceptible to amiloride (Fig. 2D), and indicate correlation between sensitivity to amiloride and ErbB3 localization. However, in both C4-2 cells (Fig. 2E), as well as in 22Rv1 (Fig. 2F), amiloride treatment increased ErbB3 localization to the plasma membrane– as indicated by co-localization with NHE1. Taken together, these results indicate that the cytotoxic effects of amiloride correlates with its translocation from the nucleus to the cytoplasm in LNCaP and from the cytoplasm to the plasma membrane in the CRPC lines.
Fig. 2
Amiloride does not promote ErbB3 translocation from the nucleus to the cytoplasm and the plasma membrane in CRPC cells (A) Hormone-insensitive C4-2 or (C) the unrelated cell line 22Rv1 cells were also treated with varying concentrations of amiloride for 72 h before being subjected to viability assays or lysed and fractionated as previously described. For all viability assays, results were obtained from triplicate experiments. Error bars represent standard deviation. Tables show p-values with respect to DMSO for each tested cell line. All densitometry was performed using ImageJ. C = cytoplasmic and N = nuclear. (B, D) Cells were subjected to viability assays using the stated concentrations of amiloride. p-values are calculated with respect to DMSO. (E, F) C4-2 and 22Rv1 cells were treated with IF-specific antibodies to NHE-1 (prototypical target of amiloride) and imaged as described in previous figure legends
The tyrosine kinase HER2, that regulates the activation of downstream targets, is itself activated by the ErbB3 ligand HRG1We next investigated the effects of amiloride on the phosphorylation status of the EGFR family and their prominent downstream targets, considered a measure of activation of these proteins. Transcript levels of any of the ErbB family members were largely unchanged across cell lines, except for EGFR mRNA in 22Rv1 cells which increased with 75µM amiloride (Supp. Figure 1A). We previously showed that the ErbB3 ligand HRG1, but not the EGFR ligand EGF, stimulated ErbB3 nuclear translocation [24]. Hence, we probed the effect of both EGF and HRG1 on ErbB receptor phosphorylation that corresponded to their activation in the presence or absence of 75µM amiloride. EGFR activation was determined by its phosphorylation at Y1068, a Grb2 binding site. EGF, but not HRG1, induced EGFR phosphorylation, and this effect was not altered by amiloride in any of the cell lines investigated (Fig. 3A). In LNCaP cells, HER2 phosphorylation at Y1248 was, however, stimulated by EGF under control conditions, whereas HRG1 activated it only upon amiloride treatment. In the CRPC lines, however, it was stimulated by both EGF as well as HRG1, but amiloride did not affect this distribution (Fig. 3A). On the other hand, in all three cell lines, ErbB3 phosphorylation at Y1289 was stimulated by HRG1 but not EGF.
Fig. 3
Differential activation and dimerization of ErbB family members and their downstream targets in HSPC and CRPC cells with high concentrations of amiloride (A) HSPC (LNCaP) and CRPC (C4-2, 22Rv1) cells were treated for 72 h with 75µM amiloride dissolved in 100% sterile DMSO and stimulated with PBS, EGF or HRG for 15 min prior to collection to observe activation of ErbB family members and their downstream targets. Cells were lysed in denaturing lysis buffer before being analysed by immunoblotting. 25 µg of protein were loaded per lane. Hsp90 was used as a loading control. (B,C) LNCaP cells (HSPC) or (D,E) C4-2 cells (CRPC) or (F, G) 22Rv1 cells (unrelated CRPC cells) were treated with 75µM amiloride or 100% DMSO (0.1% v/v) for 72 h and stimulated with PBS, EGF or HRG for 15 min to activate ErbB family dimers just prior to collection. 400ug of whole cell lysate were used in each pulldown lane. Mouse IgG antibody was used as an isotype control. Amiloride increases ErbB3-HER2 dimers and stabilizes ErbB3-EGFR dimers
Next, we investigated the downstream targets of the ErbB receptors. Both ERK and Akt are activated by all four receptors, but ErbB3 is more likely to activate Akt, given that it has six PI3Kp85 binding sites (PI3K being upstream of Akt), compared to EGFR and ErbB2 [12, 13, 15]. Similar to ErbB3 activation, Akt phosphorylation was induced by HRG1 mostly in the presence of amiloride in LNCaP cells, whereas in C4-2 and 22Rv1 cells, it was activated both in the presence and absence of amiloride (Supp. Figure 1B). In contrast, ERK was phosphorylated mostly in the same pattern as HER2 phosphorylation (Supp. Figure 1B). Taken together, these results suggest that EGF activates EGFR, HRG1 activates ErbB3, but both EGF and HRG activate HER2 and downstream targets. Since HER2 does not have a known ligand, its activation indicates heterodimerization with EGFR or ErbB3.
Amiloride selectively promotes HER2/ErbB3 heterodimerization and inhibits HRG1-induced EGFR/ErbB3 heterodimersWe next investigated whether amiloride-mediated activation of downstream EGFR family targets could be explained by receptor dimerization patterns. In LNCaP cells, HRG1, but not EGF, stimulated EGFR/ErbB3 dimerization, that was suppressed by amiloride (Fig. 3B) whereas amiloride significantly increased HER2/ErbB3 dimerization independent of ligand binding (Fig. 3B). In contrast, EGFR/HER2 dimers were diminished upon HRG1 stimulation independent of the presence of amiloride (Fig. 3C), suggesting that under control conditions, HRG1 enabled realignment of EGFR from HER2 binding to ErbB3 binding, whereas upon amiloride treatment, the EGFR/ErbB3 dimers were diminished, while HER2/ErbB3 dimers were stabilized (Fig. 3C). This is likely due to the decrease in EGFR protein levels with 75 µM treatment observed in Fig. 1A in these cells. Loss of EGFR/ErbB3 levels upon high dose amiloride treatment likely explains why at even higher doses (100 µM), nuclear ErbB3 is restored despite increased EGFR levels at that dose.
Similar to LNCaP, C4-2 cells also saw increased HRG-activated EGFR-ErbB3 dimers that were suppressed by amiloride treatment (Fig. 3D) but in these cells, there was a decrease in HER2/EGFR dimers with amiloride treatment both in the presence of EGF and HRG1 (Fig. 3D). In these cells, HER2/ErbB3 was reduced by EGF treatment, but not by HRG1 (Fig. 3E) suggesting realignment of HER2 from ErbB3 to EGFR upon EGF stimulation, that was reversed by amiloride. Amiloride promoted co-localization of HER2 with ErbB3 in these cells is better illustrated by immunofluorescent imaging (Supp. Figure 2).
In 22Rv1, again, HRG1 induced EGFR/ErbB3 dimerization, that was suppressed by amiloride (Fig. 3F), which also stabilized HER2/ErbB3 dimerization (Fig. 3F), but showed no significant change in EGFR/HER2 dimers (Fig. 3G), perhaps due to the very high levels of EGFR expression in these cells (Supp. Figure 1A), in contrast to LNCaP, where amiloride suppresses EGFR protein levels, perhaps due to a post-translational modification step. Thus, a comparison of HSPC LNCaP cells with CRPC C4-2 and 22Rv1 cells shows that in both HSPC and CRPC lines, HRG1 realigned EGFR from HER2 to ErbB3, whereas in CRPC lines, EGF realigned HER2 from ErbB3 to EGFR. In all three lines, HRG1 stimulated EGFR/ErbB3 dimerization, whereas amiloride prevented this effect, while promoting HER2/ErbB3 dimers.
Effect of HER2 receptor tyrosine kinase on ameliorating C4-2 cell viability is enhanced by amiloride treatmentWe next investigated whether silencing the receptors would decrease cell viability. We used EGFR-, HER2- or ErbB3-specific silencing RNA (siRNA) sequences at a concentration of 10 pM per treatment condition to determine whether EGFR family receptors were involved in decreasing cell viability in response to amiloride treatment. The efficacy and specificity of EGFR family silencing has been previously assessed by us [23]. We used CRPC C4-2 cells to test the effect of EGFR family on mediation of the effects of this drug. C4-2 cells showed significant decreases in viability with EGFR and ErbB3 siRNAs individually (approximately 75% decrease in viability using each siRNA) (p = 0.0015) (Fig. 4A). As before, amiloride significantly inhibited C4-2 cell growth, but knockdown of EGFR in the amiloride treated cells had no further effect, although ErbB3 knockdown slightly decreased viability (p = 0.0321) (Fig. 4A). However, knockdown of HER2 in C4-2 cells reduced viability by about 30% (p < 0.0001), whereas in amiloride treated cells, the same knockdown reduced viability by an additional 10% (p = 0.0041) (Fig. 4B). The efficacy of the siRNAs to EGFR, HER2, ErbB3 in C4-2 cells is shown in Fig. 4C. Thus, treatment with amiloride increased sensitivity to HER2 knockdown, and explains the important role of this receptor in CRPC cell viability.
Fig. 4
Amiloride efficacy is enhanced by EGFR knockdown in HSPC cells and by HER2 knockdown in CRPC cells. (A) CRPC C4-2 cells were transfected with control (CT) or EGFR or ErbB3 siRNA or (B) HER2 siRNA and treated with or without 75µM amiloride before being analysed for changes in viability with the MTT assay. Error bars represent standard deviation. Experiments were performed in triplicate. (C) Whole cell immunoblot for siRNA efficacy. 20 µg of protein were loaded per lane. Tubulin was used as a loading control. (D) 22Rv1 cells (CRPC) were transiently transfected with empty vector (EV), EGFR (B1), ErbB2 (B2), ErbB3 (B3) or mutant ErbB3 (mB3) as previously described by us in detail [23, 45]. Cells were collected, fractionated and analysed by immunoblot as described in previous figure legends. (E) ErbB3 overexpression was visualized microscopically in 22Rv1 cells using the reagents and procedures as described in previous figure legends
Overexpression of ErbB3 promotes its nuclear localizationSince HRG1 appears to promote dimerization of HER2 with ErbB3, and the impact of nuclear ErbB3 on this dimerization, we next investigated whether overexpression of ErbB3 would affect its nuclear localization. EGFR, HER2 and HER3 was overexpressed in 22Rv1 cells as indicated (Fig. 4D). We used 22Rv1 cells as these cells– unlike LNCaP which showed strong nuclear staining or C4-2 cells, which showed almost no nuclear staining, demonstrated small amounts of nuclear ErbB3. Overexpression of HER3/ErbB3 caused significant increase in nuclear ErbB3 levels (Fig. 4D), indicating that de novo ErbB3 expression results in nuclear localization, whereas it then has to be transported to the cytoplasmic compartment. These observations are supported by immunofluorescent pictures showing that overexpression of ErbB3 in 22Rv1 cells resulted in nuclear expression (Fig. 4E). To determine whether phosphorylation of ErbB3 at major tyrosine kinase sites (Y1222, Y1289, and Y1328) contributed to its nuclear localization, we mutated these sites to alanine and overexpressed the mutant plasmid (mHER3). There was no change in nuclear ErbB3 expression indicating that phosphorylation of ErbB3 did not contribute to its nuclear localization. Taken together, the results shown so far indicate that (i) EGFR and HER2 were mainly plasma membrane located in PCa cells while ErbB3 may localize on the membrane, the cytoplasm or the nucleus; (ii) amiloride promoted ErbB3 translocation from the nucleus to the cytoplasm and the cytoplasm to the plasma membrane; (iii) as a result, amiloride enhanced HER2/ErbB3 dimerization, (iv) HER2 was activated by dimerization with ErbB3 and (v) activated HER2 enhanced downstream signaling and cell proliferation.
Amiloride enhances the sensitivity of HSPC cells to lapatinibWe have previously shown that physiological concentrations of the FDA-approved reversible HER2 kinase inhibitor lapatinib were ineffective in inhibiting the growth of PCa cells [23]. Based on the data above, we hypothesized that the addition of amiloride to lapatinib would enhance cell killing efficacy, at physiological doses of both drugs. We therefore investigated the effects of 10 µM amiloride, the least dose usually used in cell lines [47] on sensitizing PCa cells to lapatinib.
We previously demonstrated that 2 µM lapatinib was the least dose that was effective in PCa cells [23]. We investigated the viability of LNCaP cells treated with increasing concentrations of lapatinib and established an IC50 = 3 µM (Fig. 5A); hence we used 2 µM as a suboptimal dose, low dose amiloride (10µM) or a combination of 2µM lapatinib and 10µM amiloride (‘2µM Lap + 10µM Amil’) to determine whether amiloride enhanced the efficacy of lapatinib in these cells. The viability of LNCaP cells decreased only 36% with 2µM lapatinib (p = 0.0925) compared to 90.5% at the highest concentration of lapatinib tested (10µM, p = 0.0192). 10µM amiloride individually produced a reduction of 63.2% (p = 0.0252). When 2 µM lapatinib was combined with 10 µM amiloride, the resulting decrease in viability was 74.2% (p = 0.0259) which was comparable to 10 µM lapatinib alone (p = 0.0091) (Fig. 5B).
Fig. 5
Amiloride enhances the sensitivity of HSPC cells to low concentrations of lapatinib (A, B) LNCaP cells were treated with 1–10µM lapatinib and assayed for viability. Lapatinib and amiloride were both dissolved in 100% DMSO. In a head-to-head comparison of lapatinib and amiloride, the combination was additive when cell viability was assayed with the MTT reagent. Experiments were done in triplicate. Error bars represent standard deviation. (C) Cells were treated with varying concentrations of lapatinib for 72 h before being collected and lysed into cytoplasmic and nuclear fractions as described in earlier figure legends. (D) Cells were treated with 2µM lapatinib, 10µM amiloride or a combination of the two for 72 h before being collected, fixed and processed for indirect immunofluorescent microscopy using immunofluorescent-specific antibodies to the C- and N- termini of ErbB3 (‘CTD’ and ‘NTD’ respectively) as previously described. Scale bars = 7.5 μm. Co-administration of lapatinib and amiloride increases the accumulation of ErbB3 compared to 2µM lapatinib alone. (E) LNCaP cells were treated for 72 h with lapatinib, amiloride or the combination or 100% sterile DMSO and stimulated with PBS, EGF or HRG for 15 min prior to collection to observe activation of ErbB family members and their downstream targets. Cells were lysed in denaturing lysis buffer before being analysed by immunoblotting. 25 µg of protein were loaded per lane. Tubulin was used as a loading control
When the expression and localization of total protein levels of EGFR family members were analyzed, little change was seen in EGFR or HER2 with lapatinib (Fig. 5C). In contrast, ErbB3 nuclear localization decreased in a dose-dependent manner from 0–10µM lapatinib (Fig. 5C). At 2 µM lapatinib, though, sufficient levels of ErbB3 remained in the nucleus. To overcome this, immunofluorescent analysis of ErbB3 localization using an anti-C-terminal (CTD) and an anti-N-terminal (NTD) ErbB3 antibody depicted nuclear localization of ErbB3 under control conditions (Fig. 5D). 2 µM lapatinib or 10µM amiloride did not disturb this pattern (Fig. 5D). However, immunofluorescent imaging showed that with 2µM lapatinib treatment in the presence of amiloride, ErbB3 localization was cleared from the nucleoplasm. An immunoblot analysis of the signaling cascades under all the treatment conditions revealed that EGFR underwent phosphorylation (or activation) with EGF treatment, ErbB3 was phosphorylated with HRG1 and HER2 with both (Fig. 5E). With 10 µM amiloride, unlike 75 µM, there was no change in EGF-induced EGFR and HER2 phosphorylation or in HRG1-induced ErbB3 phosphorylation, but HRG1-induced HER2 phosphorylation was severely affected (Fig. 5E). Significantly, lapatinib treatment, with or without amiloride, abrogated EGFR and HER2 phosphorylation in LNCaP cells but ERK phosphorylation was eliminated only with the combination (Fig. 5E).
In contrast to LNCaP cells, in 22Rv1 cells, which had very low baseline levels of nuclear ErbB3, this RTK remained cytoplasmic with lapatinib treatment, as well as with amiloride combinations, similar to EGFR and HER2 (Fig. 6A). These cells did however show a dose-dependent decrease in viability from 0–10µM lapatinib, with a 49% decrease with 5 µM lapatinib (p = 0.0157) and a 96% decrease at 10 µM lapatinib (p = 0.0075) with a resultant IC50 of 5.108 µM (Fig. 6B). However, the 10 µM intratumoral dose will not be physiologically relevant since achievement of that dose will put patients in conditions that will subject them to various adverse events prior to achieving that dose. To determine whether the more physiological dose of 2 µM can be enhanced by the addition of amiloride, we tested the combination of the two drugs in 22Rv1 cells (Fig. 6C). As before, 10 µM amiloride (that will not cause hyperkalemia) had by itself a small effect on the viability of 22Rv1 cells (p = 0.0007); however– the combination of 2 µM lapatinib and 10 µM amiloride reduced 22Rv1 viability by 49.9% (p = 0.0012), and the combinatorial effect was more significant in this cell line (than in LNCaP cells) in comparison to either lapatinib alone (p = 0.0131) or amiloride (p = 0.0164). Although there was no significant effect on AR transcriptional activity at these drug concentrations (Supplementary Fig. 3A), lapatinib significantly suppressed the activation of the RTKs and their downstream targets, with or without the presence of amiloride (Fig. 6D). Significantly, as in LNCaP cells, ERK phosphorylation was significantly decreased in HRG1-stimulated 22Rv1 cells upon combinatorial treatment, compared to lapatinib alone (Fig. 6D). while amiloride but not lapatinib eliminated any nuclear ErbB3 that still may remain in these cells (Supplementary Fig. 3B). Lapatinib (2 µM) and amiloride (10 µM) together showed a combinatorial effect when plotted against increasing doses of each drug individually (Supplementary Fig. 3C).
Fig. 6
Amiloride enhances the sensitivity of HSPC and CRPC cell lines to low concentrations of lapatinib. (A) 22Rv1 cells were treated with varying concentrations of lapatinib, 10µM amiloride or a combination of the two for 72 h before being collected and lysed into cytoplasmic and nuclear fractions as described in earlier figure legends. Co-administration of lapatinib and amiloride does not increase the accumulation of ErbB3 in the cytoplasmic fraction. (B) 22Rv1 cells were treated with 1–10µM lapatinib and assayed for viability. Lapatinib was dissolved in 100% DMSO. Experiments were done in triplicate. Error bars represent standard deviation. (C) In a head-to-head comparison of lapatinib and amiloride, the combination was additive when cell viability was assayed with the MTT reagent. Experiments were done in triplicate. Error bars represent standard deviation. Coloured dotted line estimates 50% viability. Table shows p-values with respect to FBS DMSO. (D) 22Rv1 (CRPC) cells were treated for 72 h with lapatinib, amiloride or the combination or 100% sterile DMSO and stimulated with PBS, EGF or HRG for 15 min prior to collection to observe activation of ErbB family members and their downstream targets. Cells were lysed in denaturing lysis buffer before being analysed by immunoblotting. 25 µg of protein were loaded per lane. Tubulin was used as a loading control. Tables show p-values with respect to PBS DMSO in each cell line tested. (E) PC-346 C cells were treated with 10–60 µM of amiloride and assayed for viability. Amiloride was dissolved in 100% DMSO. Experiments were done in triplicate. Error bars represent standard deviation. (F) PC-346 C cells were treated with 1–10µM lapatinib and assayed for viability. Lapatinib was dissolved in 100% DMSO. Experiments were done in triplicate. Error bars represent standard deviation. (G) PC-346 C cells were treated with lapatinib, amiloride or the combination, which is shown to be additive when cell viability was assayed with the MTT reagent. Experiments were done in triplicate. Error bars represent standard deviation. (H) PC-346 C cells were treated with varying concentrations of lapatinib, 10µM amiloride or a combination of the two for 72 h before being collected and lysed into cytoplasmic and nuclear fractions as described in earlier figure legends. (I) Cells were treated with 2µM lapatinib, 10µM amiloride or a combination of the two for 72 h before being collected, fixed and processed for indirect immunofluorescent microscopy using immunofluorescent-specific antibodies to the C-terminal domain of ErbB3 as previously described. Yellow boxes (inset) and bold yellow arrow depict area of negligible ErbB3 nuclear staining. Scale bars = 7.5 μm
Thus far, we have used three cell lines– that are sensitive to amiloride (IC50 in the range 20–40 µM), however, we then investigated the effect of the combination on PC-346 C cells, previously reported by us [12] that is more resistant to amiloride [IC50 = 67.38 µM (55.26 µM -108.3 µM)] (Fig. 6E). These cells express wild type AR at very low levels and are considered to be hormone sensitive since they are inhibited by flutamide [48]. We therefore tested the effect of the amiloride-lapatinib combination on PC-346 C cells, which are, however, very sensitive to lapatinib [IC50 = 1.579 µM (1.203 µM -2.001 µM)] (Fig. 6F). While 10 µM amiloride had no effect on PC-346 C cells, 2 µM lapatinib caused a 68% decrease in viability (p = 0.0003) (Fig. 6F). The combination of lapatinib and amiloride caused an additional 32.5% decrease in viability (p = 0.0440 compared to lapatinib alone) (Fig. 6G), indicating a combinatorial effect even in these amiloride-resistant cells.
To determine whether the mechanism by which the combination works in a second hormone-sensitive PCa cell line PC-346 C [49] is similar to that in LNCaP cells, we tested the effects of these treatments on EGFR, HER2 and ErbB3. Like the other lines, EGFR and HER2 was mostly cytoplasmic, and remained so, irrespective of the treatment. ErbB3 was partly nuclear, and the nuclear expression was enhanced by amiloride treatment, which explains its resistance to this drug (Fig. 6G). In contrast, lapatinib alone did not affect ErbB3 nuclear levels, but in the presence of amiloride, significantly reduced ErbB3 nuclear localization further, explaining the additive effect on cell viability (Fig. 6H). This is reinforced by immunofluorescent imaging showing that the combination of lapatinib and amiloride removes the levels of nuclear ErbB3 (Fig. 6I). Taken together, in hormone sensitive cells, the presence of nuclear ErbB3 induces resistance to cell death, whereas treatment with lapatinib and amiloride reduces ErbB3 nuclear localization and reduces viability.
Low dose amiloride and lapatinib combine to induce apoptosis in HSPC cellsThe goal of cancer treatment is to ensure that all malignant cells are dead, not dormant. However, the changes in cell viability that we have conducted thus far could be due to an increase in apoptosis, or the onset of various mechanisms that may have led to cellular quiescence. Hence, we conducted cell death analyses to ascertain the mechanism causing the consistent decreases in viability seen in all 3 cell lines with the combination of low dose lapatinib and amiloride. Flow cytometry was employed using DNA-bound propidium iodide (PI) as a necrosis marker and cell surface expression of Annexin V using the Annexin V-Allophycocyanin (APC) conjugate as a marker of apoptosis. LNCaP cells showed no significant change in early apoptosis (‘APC’) or late apoptosis (apoptosis with necrosis, ‘PI + APC’) with 2µM lapatinib and a slight decrease in early apoptosis (-39.6%) with 10µM amiloride (p = 0.023) (Fig. 7A). The combination of 2µM Lap + 10µM amiloride produced a sharp increase in the percentage of cells undergoing early apoptosis (2.7-fold, p = 0.0061) and this was increased further when amiloride was used at 75µM (4.42-fold, p < 0.0001), although no change in the fraction of cells in late apoptosis was noted (Fig. 7A). In contrast, C4-2 cells exhibited an increase in necrotic cells only when 2 µM lapatinib + 75µM amiloride were used (2.26-fold, p = 0.0371) which may indicate toxicity, rather than programmed cell death, while a combination of 2 µM lapatinib + 10µM amiloride actually resulted in a 65% decrease in apoptosis (p = 0.0193) (Fig. 7B). Similar to C4-2 cells, 22Rv1 cells displayed no change in apoptosis when exposed to 2µM lapatinib either alone or in combination with 10 µM or 75 µM amiloride, and significantly reduced cell death with 2µM lapatinib in combination with both 10µM and 75µM amiloride (Fig. 7C). Representative raw readings for these data are provided in supplementary information (Supp. Figs. 4, 5 and 6). Thus, in LNCaP cells, the decrease in cell viability with the combination of 2 µM lapatinib + 75µM amiloride observed is likely due to an increase in apoptosis while that in C4-2 cells, any change in viability is likely caused by increase in toxicity, and no substantial effects of the combination was observed in 22RV1 cells. Taken together, this indicates that the combination of 2 µM lapatinib + 10µM amiloride was effective in inducing programmed cell death in HSPC LNCaP cells (higher doses may cause toxicity), but not in CRPC lines at any dose.
Fig. 7
Amiloride and lapatinib synergize to increase apoptosis in HSPC and CRPC cell lines. (A-C) HSPC and CRPC cell lines were treated with 100% DMSO, lapatinib, amiloride or the combination (in µM) for 72 h before being processed for cell death analysis using annexin V and propidium iodide staining. The percentage of cells undergoing early or late apoptosis with DMSO treatment was set to 100% and values for the various treatment conditions calculated accordingly. Experiments were performed in triplicate. Error bars represent standard deviation. (D) Schematic with proposed molecular mechanism of lapatinib-amiloride efficacy. (a) EGFR, HER2 and ErbB3 exist at the cell membrane and signal via pathways such as ERK and AKT. (b,c) ErbB3 monomers cycle between the nucleus and cell membrane. (EGFR and HER2 behave similarly but have been omitted for clarity). (d) Lapatinib is a dual-kinase TKI (tyrosine kinase inhibitor) of HER2 and EGFR dimers but will also inhibit HER2 in HER2-ErbB3 dimers. Lapatinib is unlikely able to inhibit ErbB3 if it is in the nucleus and not at the cell surface. (e) Amiloride is a macropinocytosis inhibitor that prevents internalization of ErbB3 and retains it at the cell surface. As a result, nuclear ErbB3 decreases and cytoplasmic surface ErbB3 increases. (f) Amiloride-induced ErbB3 retention enables its dimerization with HER2, enabling the formation of ErbB3-HER2 dimers which are now inhibited by the addition of low concentrations of lapatinib
留言 (0)