
記住我
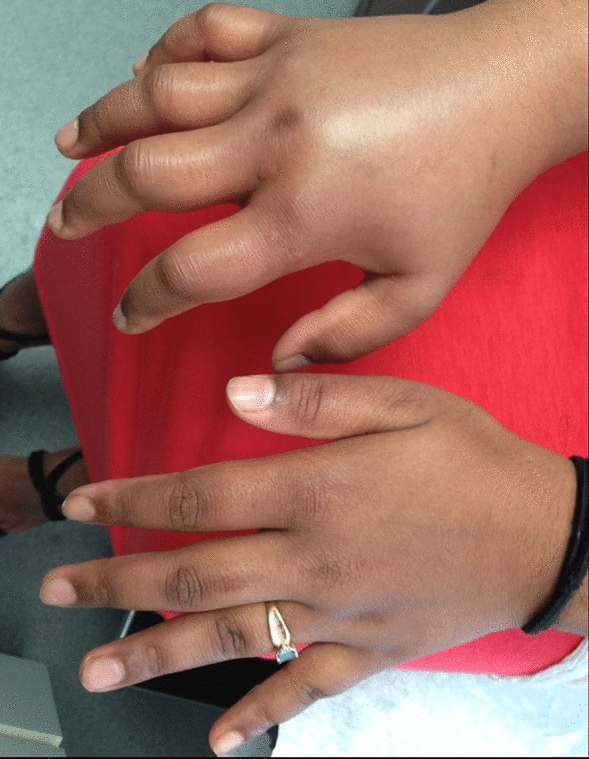

The immune system refers to a collection of cells, chemicals and processes that function to protect the body from infection and damage. At areas in contact with disease-causing organisms such as bacteria, viruses, fungi and parasites (collectively referred to as pathogens), the immune system provides ongoing active defense against infection. This activity is particularly important at body surfaces which interact with the external environment, such as the skin, airways, and gastrointestinal and reproductive tracts. In addition, the immune system protects us against cancer cells and toxins throughout the body. Beyond the structural and chemical barriers which protect us from infection, the immune system can be simplistically viewed as having two “lines of defense”, known as innate immunity and adaptive immunity. Innate immunity represents the first line of defense to an intruding pathogen. It is a defense mechanism that is used by the host immediately, or within hours, of encountering a pathogen or tissue damage. The innate immune response can be triggered by common chemical signals associated with multiple pathogens, such as structures found on bacterial or fungal cell walls. Adaptive immunity, on the other hand, is antigen-dependent and antigen-specific, which means it responds to a very precise chemical structure(s) (antigen[s]), and involves a lag time between exposure to the antigen and maximal response. The important hallmark of adaptive immunity is the capacity for memory, which enables the host to mount a more rapid and efficient immune response upon subsequent exposure to the antigen. This memory response allows the host to vigorously combat infection, and it is the basis for vaccination strategies. Innate and adaptive immunity are not mutually exclusive mechanisms of host defense, but rather are complementary and interactive, with defects in either system resulting in host vulnerability or inappropriate responses. Often, the innate immune response aids in directing a more rapid and effective acquired response to infection [1, 2].
Innate immunityInnate immunity can be viewed as comprising four types of defensive barriers: anatomic (skin and mucous membrane), physiologic (temperature, low pH and chemical mediators), endocytic and phagocytic, and inflammatory. Table 1 summarizes the non-specific host-defense mechanisms for each of these barriers. Cells and processes that are critical for effective innate immunity to pathogens that evade the initial anatomic barriers have been widely studied. Innate immunity to pathogens often relies on pattern recognition receptors (PRRs) which allow a limited range of immune cells to detect and respond rapidly to a wide range of pathogens that share common structures, known as pathogen associated molecular patterns (PAMPs). Examples of these include bacterial cell wall components, such as lipopolysaccharides (LPS), and double-stranded ribonucleic acid (RNA) produced during viral infection. In addition, the innate immune system responds to signals from dead and dying cells, allowing innate immunity to mobilize if the physical barriers that protect the body are damaged. These processes involve chemicals known as “alarmins” and damage-associated molecular patterns (DAMPs) produced in response to cell and tissue damage.
Table 1 Summary of non-specific host-defense mechanisms for barriers of innate immunityAn important function of innate immunity is the rapid recruitment of immune cells to sites of infection and inflammation through the production of cytokines and chemokines (proteins involved in immune cell–cell communication and recruitment). Cytokine production during innate immunity mobilizes defense mechanisms throughout the body while also activating local cellular responses to infection or injury. Key inflammatory cytokines released during the early response to bacterial infection are tumour necrosis factor (TNF), interleukin 1 (IL-1) and interleukin 6 (IL-6). These cytokines are critical for aiding in cell recruitment and the local inflammation and increased mucus production which is essential for clearance of many pathogens. They also contribute to the development of fever. Dysregulated production of such inflammatory cytokines, so that they are produced excessively without infection, is often associated with inflammatory or autoimmune disease, making them important therapeutic targets. Chemokines, such as CXCL8 (IL-8) and CCL2, direct the movement of critical immune cells, such as neutrophils and monocytes, into tissues to combat infection. Cytokines and chemokines produced during the innate immune response also aid in the proper development of an effective adaptive immune response by enhancing the activity of antigen-presenting cells (APCs) (discussed later) and increasing the accumulation of cells within lymph nodes draining infection sites.
The complement system is a biochemical cascade that functions to identify and opsonize (coat) bacteria and other pathogens. It renders pathogens susceptible to phagocytosis, a process by which immune cells engulf microbes and remove cell debris, and kills some pathogens and infected cells directly. The phagocytic action of the innate immune response promotes clearance of dead cells or antibody complexes and removes foreign substances present in organs, tissues, blood, and lymph. It can also activate the adaptive immune response through the mobilization and activation of APCs [1, 3].
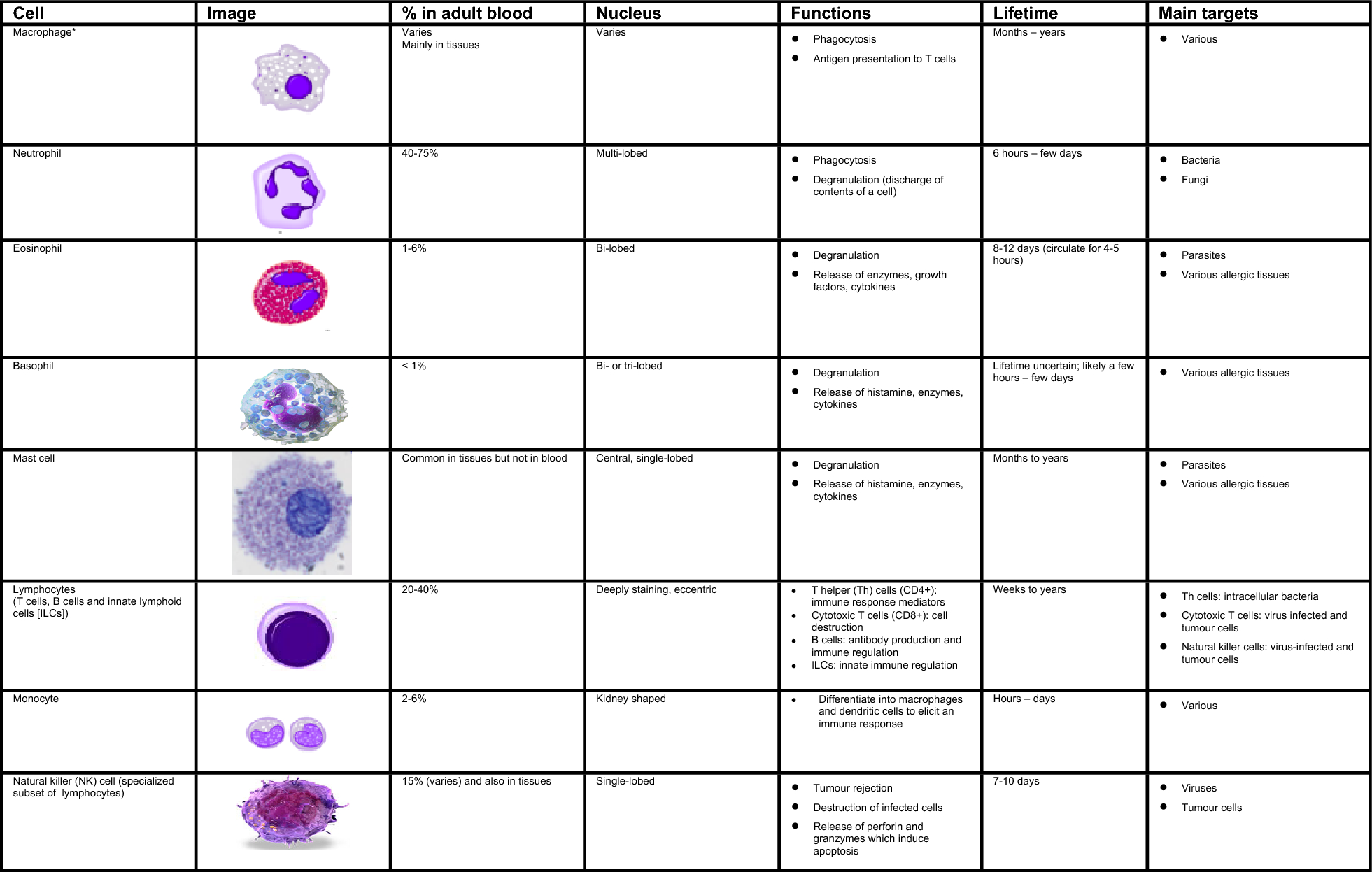
Numerous cells are involved in the innate immune response such as phagocytes (macrophages and neutrophils), dendritic cells, mast cells, basophils, eosinophils, natural killer (NK) cells, and innate lymphoid cells. The main characteristics and functions of these cells are summarized in Fig. 1 [1, 3, 4].
Fig. 1
Characteristics and function of cells involved in innate immunity [1, 3, 4]. *Include alveolar macrophages (within pulmonary alveolus), histiocytes (connective tissue), Kupffer cells (liver), microglial cells (neural tissue), epithelioid cells (granulomas), osteoclasts (bone), mesangial cells (kidney)
Phagocytes are sub-divided into two main cell types: neutrophils and macrophages. Both cell types share a similar function: to engulf (phagocytose) microbes and kill them through multiple bactericidal pathways. In addition to their phagocytic properties, neutrophils contain granules and enzyme pathways that assist in the elimination of pathogenic microbes. Unlike neutrophils, which are short-lived cells mobilized rapidly to sites of infection, macrophages are long-lived cells that patrol the body and remove pathogens, dead cells, and debris. These cells not only play a role in phagocytosis, but are also involved in antigen presentation to T cells [1].
Dendritic cells also phagocytose and function as APCs, initiating the acquired immune response and acting as important linking cells between innate and adaptive immunity. Mast cells and basophils share many features with each other, and both are instrumental in the initiation of acute inflammatory responses, such as those seen in allergy and asthma. Mast cells also have important functions as immune “sentinel cells” and are early producers of cytokines in response to infection or injury. Unlike mast cells, which generally reside in the connective tissue surrounding blood vessels and are particularly common at mucosal surfaces, basophils generally reside in the circulation and are recruited to tissues following infection or during chronic allergic disease. Eosinophils are granulocytes that possess phagocytic properties and play an important role in the destruction of parasites that are often too large to be phagocytosed through toxic granule product release. Along with mast cells and basophils, they are also associated with allergy and asthma. NK cells play a major role in the rejection of tumours and the destruction of cells infected by viruses. Destruction of infected cells is achieved through the release of perforins and granzymes (proteins that cause lysis of target cells) from NK-cell granules which induce programmed cell death or apoptosis [4]. NK cells are also an important source of another cytokine, interferon-gamma (IFN-γ), which helps to mobilize APCs and promote the development of effective antiviral adaptive immunity. NK cells are part of a larger group of innate lymphoid cells (ILCs); the other members of this family play a more regulatory role. Depending on their type (i.e., ILC-1, ILC-2, ILC-3), they selectively produce cytokines such as IL-4, IFN-γ, and IL-17, respectively, that help to direct the appropriate early immune response to specific pathogens and contribute to immune regulation in that tissue but do not require specific antigen stimulation to do so.
Adaptive immunityThe development of adaptive immunity is aided by the actions of the innate immune system. Adaptive immunity is critical when innate immunity is ineffective in fully eliminating infectious agents and for providing a “memory” enhanced response to repeated infection (e.g., to seasonal viruses like influenza or severe acute respiratory syndrome coronavirus 2 [SARS-CoV-2]). The primary functions of the adaptive immune response are the recognition of specific “non-self” antigens associated with pathogens or other threats, the generation or enhancement of immune effector pathways that can eliminate specific pathogens or pathogen-infected cells, and the development of an immunologic memory that can quickly eliminate a specific pathogen should subsequent infections occur. Adaptive immune responses are the basis for effective immunization against infectious diseases. The cells of the adaptive immune system include antigen-specific T cells, which are activated to proliferate through the action of APCs and have several roles in combatting infections and cancer, as well as B cells which differentiate into plasma cells to produce antibodies.
T cells and APCsT cells derive from hematopoietic stem cells in bone marrow and, following migration, mature in the thymus. These cells express a series of unique antigen-binding receptors on their membrane, known as T-cell receptors (TCR). Each T cell expresses a single type of TCR, which binds to a particular unique chemical structure on an antigen. There are several thousand unique TCR on T cells in every individual. These T cells have the capacity to rapidly divide, proliferate, and differentiate into a T cell type(s) with specific roles in immunity. These roles include regulation of the immune system, promotion of antibody production by B cells and direct killing of virally infected or cancer cells. T cells divide and proliferate only if they receive the appropriate signals through the action of APCs (usually dendritic cells, but sometimes macrophages, B cells or structural cells) to recognize a specific antigen.
The surfaces of APCs express a group of proteins known as the major histocompatibility complex (MHC). MHC are classified as either class I (also termed human leukocyte antigen [HLA] A, B and C) which are found on all nucleated cells, or class II (also termed HLA-DP, -DQ and -DR) which are found only on certain cells of the immune system, including macrophages, dendritic cells, and B cells. Class I MHC molecules present endogenous (intracellular) peptides, such as viral peptides in virally infected cells, while class II molecules on APCs present exogenous (extracellular) peptides, such as those from many bacteria to T cells. These MHC proteins display fragments of antigens (peptides) when a cell is infected with an intracellular pathogen, such as a virus, or has phagocytosed foreign proteins or organisms [2, 3, 5].
T cells have a wide range of unique TCRs which can bind to specific foreign peptides. During the development of the immune system, T cells that would react to antigens normally found in our body are largely eliminated, and other immune mechanisms reduce T-cell responses to items usually found in our environment, such as food. T cells are activated when they encounter an APC that has digested an antigen and is displaying the correct antigen fragments (peptides) bound to its MHC molecules to bind to that T cell’s specific TCR. The opportunities for the correct T cells to be in contact with an APC carrying the appropriate peptide MHC complex are increased by the circulation of T cells throughout the body (via the lymphatic system and blood stream) and their accumulation (together with APCs) in lymph nodes, a process that is enhanced by lymph node swelling initiated by the innate immune response. The MHC-antigen complex activates the TCR and the T cell secretes cytokines which further control the immune response. This antigen presentation process, in combination with signals provided by local cytokines, stimulates T cells to differentiate primarily into cytotoxic T cells (CD8 + cells) or T-helper (Th) cells (CD4 + cells) (see Fig. 2).
Fig. 2
Fundamentals of the adaptive immune response. Figure created using Biorender
CD8 + cytotoxic T cells are primarily involved in the destruction of cells infected by foreign agents, such as viruses or intracellular bacteria, as well as the killing of some types of tumour cells. These cells are activated by the interaction of their TCR with peptide bound to MHC class I molecules, which can be found on most cells. Clonal expansion of cytotoxic T cells produces effector cells which release substances that induce apoptosis of target cells. Upon resolution of the infection, most effector cells die and are cleared by phagocytes. However, a few of these cells are retained as memory cells that can quickly differentiate into effector cells upon subsequent encounters with the same antigen or pathogen [3, 5].
CD4 + T helper (Th) cells play an important role in establishing and maximizing the immune response. These cells have no cytotoxic or phagocytic activity and cannot directly kill infected cells or clear pathogens. Instead, they promote the immune response by directing other cells to perform these tasks and regulate the type of immune response that develops. Th cells are activated through TCR recognition of antigen bound to class II MHC molecules. Once activated, Th cells release cytokines that influence the activity of many cell types, including the APCs that activate them. The responses of CD4 cells include the production of specialized cytokines such as IL-4, IL-17 and IFN-γ that can act to enhance other arms of the immune response, such as the production of antibodies by B cells, the killing activity of NK cells, and the ability of macrophages to take up and destroy bacteria.
Several types of Th cell responses can be generated, with Th1, Th2 and Th17 being the most frequent. The Th1 response is characterized by the production of IFN-γ which activates the bactericidal activities of macrophages and enhances antiviral immunity as well as immunity to other intracellular pathogens. Th1-derived cytokines also contribute to the differentiation of B cells to make opsonizing antibodies that enhance the efficiency of phagocytes. An inappropriate Th1 response is associated with certain autoimmune diseases.
The Th2 response is characterized by the release of cytokines (IL-4, 5, and 13) which promote the development of potent antibody responses to fight extracellular infections. These include immunoglobulin E (IgE) antibody-producing B cells, as well as the development and recruitment of mast cells and eosinophils that are essential for effective responses against many parasites. In addition, they enhance the production of certain forms of IgG that aid in combatting bacterial infection. As mentioned earlier, mast cells and eosinophils are instrumental in the initiation of acute inflammatory responses, such as those seen in allergy and asthma. IgE antibodies are also associated with allergic reactions (see Table 2). Therefore, an imbalance of Th2 cytokine production is associated with the development of atopic (allergic) conditions. In contrast, Th17 cells are characterized by the production of cytokines of the IL-17 family, and are associated with ongoing inflammatory responses, particularly in chronic infection and disease. This Th17 response is regulated by several other cytokines associated with inflammation, including IL-23 from T cells and other cells. Like cytotoxic T cells, most Th cells will die upon resolution of infection, with a few remaining as Th memory cells [3, 5].
Table 2 Major functions of human Ig antibodies [6]A further subset of the CD4 + T cells, known as “regulatory T cells” (T reg), also play a role in the immune response. T reg cells limit and suppress immune responses. They also help to prevent development of autoimmune diseases by reducing immune responses to self-antigens. T reg cells may also help in the resolution of normal immune responses, as pathogens or antigens are eliminated. These cells play a critical role in the development of “immune tolerance” to certain foreign antigens, such as those found in food or expressed by commensal (helpful) bacteria in the intestine.
B cellsB cells arise from hematopoietic stem cells in the bone marrow and, following maturation, express unique antigen-specific receptors on their membrane. Unlike T cells, B cells can recognize and respond to antigens directly, without the need for APCs, through distinctive antibodies expressed on their cell surface. Each B cell expresses an antibody that recognizes only a single antigen structure, which does not have to be a protein peptide. Antibodies can also recognize lipid and carbohydrate structures produced by pathogens. The principal function of B cells is the production of antibodies against foreign antigens. This requires their further differentiation [3, 5]. Under certain circumstances, B cells can also act as APCs.
B cells undergo proliferation when activated by antigens (e.g., a particular structure on a bacterial cell surface to which they have an appropriate antigen-specific receptor). Usually with the help of signals from CD4 T cells and local cytokines, they differentiate into antibody-secreting plasma cells or memory B cells (see Fig. 2). Memory B cells are “long-lived” survivors of past infection and continue to express antigen-binding receptors. These cells can be called upon to respond quickly by producing antibodies that eliminate an antigen upon re-exposure. Plasma cells, on the other hand, are mostly shorter lived and produce large amounts of antibody that enters the circulation and tissues providing effective protection against pathogens. Given their function in antibody production, B cells play a major role in antibody-mediated immune responses (described below) as opposed to the cell-mediated immune response which is governed primarily by T cells [3, 5].
留言 (0)