MSCs were first applied in a clinical trial over a decade ago. Since then, MSCs have been used in several therapeutic approaches (Kobolak et al. 2016). There is a strong need for a universal strategy confirming the safety of MSCs, including evaluation of tumorigenicity risk, which has thus far been proven and linked mainly to pluripotent cell therapy products (ESCs and iPSCs) (Hentze et al. 2009, Yasuda et al. 2018). Although compared to pluripotent stem cells, MSCs have a significantly lower risk of oncogenic events, current methods to evaluate their safety still lack sensitivity. To address stem cell safety regulations, numerous organizations have been established, such as the Health and Environmental Sciences Institute (HESI) and the Committee for Nonclinical Safety Evaluation of Pluripotent Stem Cell-derived ProducT (FIRM-CoNCEPT) (Sato et al. 2019).
The main guidelines for culturing MSCs for therapeutic use were described by the ISCT. Additionally, two techniques were suggested to identify genetic errors: karyotyping and molecular arrays (Sato et al. 2019). Both are used to detect recurrent abnormalities but on different levels of sensitivity, which may cause more subtle mutations to be missed and thus increase the potential risk of tumorigenicity.
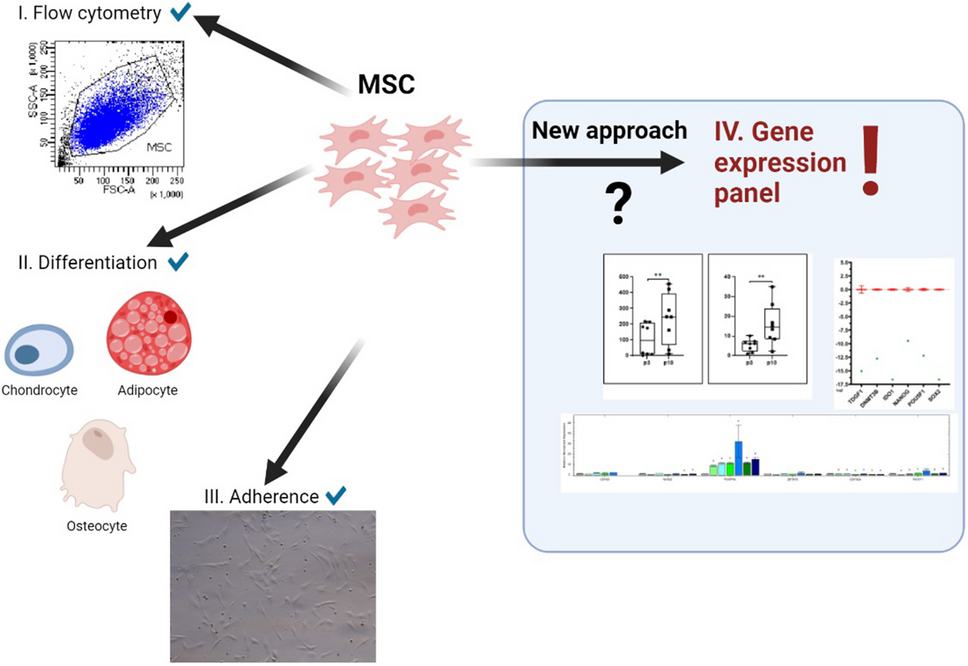
In this manuscript, we attempted to find novel ways to assess WJ-MSC tumorigenic potential. After reviewing the available literature, we selected 100 genes whose expression level was analyzed in WJ-MSC lines vs. cancer cells and iPSC lines based on a hPSC Scorecard gene panel and additional literature-based markers to define genes that help to distinguish these cell lines at the molecular level. Our WJ-MSC lines showed negligible expression of typical iPSC markers, such as TDGF1, LCK, DNMT3B, IDO1, OCT4 (POU5F1), SOX2, and NANOG. Previous studies detected the expression of ESC markers, such as SOX2, NANOG, TDGF1, and OCT4 (POU5F1) in MSCs; however, their relative expression was many times lower than that in ESCs (Riekstina et al. 2009, Heo et al. 2016, Fong et al. 2011, Gao et al. 2013). OCT4 (POU5F1), SOX2, and NANOG promote pluripotency and self-renewal through positive regulation of their genes and transcription factors (e.g., STAT3) or components of TGF-β signaling (e.g., TDGF1 and LEFTY2) (Boyer at al. 2005). Accordingly, we found that MSCs showed lower expression of LEFTY2 than iPSCs and cancer cells.
Metabolic programs are regulated by transcription factors, and in turn, metabolism controls the expression of fate regulators. ESCs and iPSCs rely mainly on glycolysis, which generates metabolites important for epigenome modifications. For example, acetyl-CoA produced by glycolysis maintains histone acetylation and pluripotency (Moussaieff et al. 2015). Overexpression of IDO1 in ES cells increases the levels of acetyl‐CoA and histone acetylation. IDO1 promotes glycolysis and pluripotency and is downregulated during the onset of hESC differentiation (Liu et al. 2019). We found that MSCs had reduced levels of IDO1 compared to iPSC and cancer cell line populations, although this was not significant. ESC renewal and differentiation are regulated by DNMT3B, a DNA methyltransferase. DNMT3B-null ESCs exhibit a disturbed mitochondrial fission and fusion balance and a switch from glycolysis to oxidative phosphorylation (Cieslar-Pobuda et al. 2020). Consistently, our MSC lines were devoid of DNMT3B expression. While iPSCs rely on glycolysis, in MSCs, there is a metabolic shift towards oxidative phosphorylation during expansion in culture in a nutrient-rich environment (Pattappa et al. 2011). Elevated glycolytic metabolism in ESCs is accompanied by increased MYC transcriptional activity and increased nuclear N-MYC and C-MYC levels (Gu et al. 2016). The MYC levels fluctuated only moderately between cancer cell, iPSC, and MSC lines. An additional aspect discriminating MSCs and iPSCs is proliferation capacity. While iPSCs maintain unlimited proliferation potential, MSCs following multiple passages acquire a senescent phenotype (Gu et al. 2006). The core transcription factors regulating pluripotency, NANOG, SOX2, and OCT4 (POU5F1), have been shown to regulate the expression of cell cycle-related genes, including positive and negative cell cycle regulators. These control pluripotency by multiple mechanisms, including feedback loops to SOX2 (Zaveri and Dhawan 2018). In our report, MSCs do not express pluripotency markers (or express them to a very limited extent); thus, MSCs cannot maintain the sustainable self-renewal stage.
We analyzed a group of genes that were upregulated in MSC lines compared to cancer cell lines, including transcription factors (ZBTB16, HAND2, and TWIST1), a cadherin regulating cell adhesion (CDH20), a receptor tyrosine kinase (PDGFR-α), and a negative cell cycle regulator (CDKN2A). These genes showed limited or no expression in iPSC lines. TWIST1 is a helix-loop-helix transcription factor. While the levels of TWIST1 were consistent among the MSC lines analyzed in this study, a previous report showed that donor-to-donor differences in TWIST1 levels correlated with MSC viability (Boregowda et al. 2015). In the same study, TWIST1 was shown to regulate gene networks controlling cell surface receptor-linked signal, transduction, cell division, and DNA replication. Intriguingly, TWIST1 regulates the expression of the p14/p16 locus in human bone marrow stromal stem cells (Cakouros et al. 2012). The authors presented a model in which high levels of TWIST1 during an early passage repress the Ink4A/Arf locus resulting from increased histone methylation by recruitment of histone methyltransferase, EZH2. As the cells senesce, the levels of TWIST1 decrease, resulting in transcriptional activation of p14/p16 (Cakouros et al. 2012). We found elevated expression of CDKN2A (coding for p14/p16) in MSCs compared to cancer cells and iPSC lines. While the levels of TWIST1 did not differ between early- and late-passage cells in our experiment, increased CDKN2A expression at later passages was observed. Transcription factors, HAND2, and its interaction with GATA4 and NKX2.5 are essential for cardiac morphogenesis. The interaction of HAND2 with TWIST1 and TWIST2 is important for limb development. HAND2 was expressed at higher levels in MSC lines than in iPSCs and cancer cell lines. HAND2 gene hypermethylation and epigenetic silencing increase the risk of endometrial cancer development (Jones et al. 2013). While we found HAND2 among genes upregulated in MSCs, HAND2 was previously not detected in human bone marrow-derived MSC lines (Riekstina et al. 2009). The difference may be attributed to the distinct tissue sources of MSCs. CDH20 (cadherin 20) has been shown to act as a cancer suppressor, and some cancer cell lines (e.g., melanoma cell line) lack the expression of CDH20 (Moore et al. 2004). CDH20 interacts with β-catenin, causing a reduction in the phosphorylation and nuclear translocation of Smad2/3, thus suppressing transforming growth factor-β (TGF-β) signaling (Li et al. 2020) and partially blocking the epithelial to mesenchymal transition (EMT) by downregulating the Snail/Smad2/3 pathway. We show that MSCs express CDH20, thus presenting another security switch for cell cycle control and repression of potential oncogenic transformation. Comparison of gene expression in the custom-designed assay showed that the expression of B4GALNT1, SOX9, and HER2 increased during later passages. B4GALNT1 encodes β−1,4-N-acetyl-galactosaminyltransferase 1, a key enzyme in generating gangliosides GM2/GD2, and plays a role in regulating cell adhesion (Martinez et al. 2007). The expression of B4GALNT1 in our MSC lines also increased during later passages. Previous studies reported that B4GALNT1 is expressed by MSCs derived from different tissues (Heo et al. 2016, Martinez et al. 2007). A study showed that the SOX9 transcription factor is expressed at similar levels in bone marrow-derived MSCs as in ESCs (Riekstina et al. 2009). Despite its role in chondrogenesis, it plays an important role in cell survival and proliferation of MSCs by controlling the levels of cell cycle regulators (Stöck et al. 2013). HER2 is a receptor tyrosine-protein kinase that regulates cell signaling, and it is a known cancer marker. It is also present in bone marrow MSCs, and its expression is enriched in a gene cluster related to proliferation (Khong et al. 2019).
Finally, we pinpointed significant overexpression of receptor tyrosine kinase alpha (PDGRF-α), a protein-coding gene responsible for mediating important cellular processes, such as proliferation, cell growth, and differentiation (Farahani and Xaymardan 2015), by activation of multiple downstream pathways, including VEGF, PI3K/Akt, and PLCγ/PKC. Some reports state that abundant amounts of PDGFR-α are a characteristic feature of undifferentiated MSCs, in which it acts as a differentiation inhibiting factor and plays critical functions during embryonic development (Ball et al. 2007). Thanks to the important crosstalk between VEGF-A and PDGFR-α, PDGFR-α is responsible for MSC migration and plays a role in neovascularization (Farooqi and Siddik 2015). Although it may be implicated during tumorigenesis due to the promotion of vasculogenesis, overexpression of PDGRF-α in mature tissues is now suggested as a novel marker of tumors. Based on Farooqi et al. (2015), PDGFR-α is repressed by p53 to regulate both neovascularization and cellular proliferation.
Using String software, we analyzed interaction networks for all panels of protein-coding markers. In this approach, the String analysis was inevitably biased, due to the use of a randomized but preselected pool of genes with overrepresented groups of genes concerning cell development and differentiation. Nevertheless, the analysis showed that panels shared a common regulator, p53. The tumor suppressor p53 regulates cellular processes, such as cell cycle control, differentiation, and DNA repair. Mutations in the p53 gene lead to genome instability and alterations in cell proliferation, the most common cancer-related genetic defect. MSCs are sensitive to mutations in the p53 gene and negative cell cycle regulator p21 (CDKN1A) (Farahani and Xaymardan 2015).
Loss of p53 expression in the absence of p21 expression in MSCs resulted in the formation of fibrosarcomas (Farahani and Xaymardan 2015), and it might have a connection with PDGFR-α-dependent tumorigenesis. Accordingly, it was shown that normal p53 expression is indispensable for MSC integrity (Boregowda et al. 2018). CDKN2A, coding for p16INK4A and p14ARF, acts as a negative regulator of the proliferation of normal cells. p16INK4A interacts with cyclin-dependent kinases 4 and 6 (CDK4 and CDK6). p14ARF blocks the activity of MDM2, an E3 ubiquitin ligase that regulates the activity and stability of p53. Because p53 is important for iPSC self-renewal (Abdelalim and Tooyama 2012), elevated expression of CDKN2A may be a marker that distinguishes MSCs from iPSCs and cancer cell lines. Accordingly, CDKN2A expression increased during later passages of MSCs, again suggesting that MSCs do not maintain the ability to proliferate indefinitely. Thus, the elevated expression of negative cell cycle regulators such as CDKN2A may be a key determinant of the limited proliferation potency of MSC lines.
In this study, researchers compared 98 genes for their expression in MSC together with cancer lines of different embryonic origin and IPS lines. The goal was to find genes that distinguish MSCs when compared to other cell populations (cancer cell lines and iPSCs) which would allow substantial improvements to currently used methods of identifying the population of MSC cells. On the other hand, detection of characteristic markers distinguishing both pluripotent and cancer cells would significantly improve the safety of preparations based on MSC due to their negative selection. In this study, researchers managed to detect over a dozen such genes, which may be the basis for the further development of the identification panels. Among the examined genes, MSCs showed the greatest difference in expression compared to other lines in such genes as PDGRF-a, CDH20, and CDKN2A. IPS lines were characterized by significant overexpression in the case of genes generally considered as pluripotency markers such as DNMT3B, IDO1, LCK, and TDGF1 but also GDF3 or ELAVL3. In addition, although it was not in the spectrum of main interest, a set of potentially usable genes for distinguishing characteristically MSC lines from cancer lines was detected, including those of different embryonic origins such as FUT3, NOS2, COLEC10, TRPM8, CLDN1, MYO3B, CDH5, TM4SF1, SST, PTHLH, and NPPB. However, the type of analysis presented here leads to the loss of information about the specific differences between MSC and cancer cell line expression. Selected genes, when averaged, show the most prominent tumorigenicity vs. MSC markers that are stable regardless of the tumor type. Due to the differences, these genes are most probably related to the mechanisms of uncontrolled proliferation, which is a key feature of tumors. Thus, there is a high chance that the cancer cell markers selected based on presented approach will be highlighted similarly in any other randomly selected cancer.
We proposed the use of selected genes to create a gene panel to characterize MSC populations and exclude their overlapping with cancer cell line expression. This could further increase confidence in patients, physicians, and manufacturers, concerning the quality and safety of products based on MSC. Diversity of MSC gene expression profile depends on the culture medium and isolation site. In this study, we aimed to ensure that cells did not exhibit gene expression similar to cancers, with specific for MSC gene expression within the range.
A fine-tuning of the designed tool requires more samples analyzed also from other than umbilical blood cord source of MSC, to create a fully reliable assay.




留言 (0)